
Scopoletin: Anticancer potential and mechanism of action
2023-02-08 14:21:44LiaMeilawatiRitaMarletaDewiApriliaNurTasfiyatiAbdiWiraSeptamaLuciaDwiAntika
Lia Meilawati, Rita Marleta Dewi, Aprilia Nur Tasfiyati, Abdi Wira Septama, Lucia Dwi Antika✉
1Research Center for Pharmaceutical Ingredients and Traditional Medicine, National Research and Innovation Agency of Indonesia, Indonesia
2Center for Biomedical Research, National Research and Innovation Agency of Indonesia, Indonesia
3Research Center for Chemistry, National Research and Innovation Agency of Indonesia, Indonesia
ABSTRACT Cancer is considered a leading cause of death worldwide due to its high morbidity and mortality rate. Conventional treatments for cancer therapy, such as chemotherapy, and radiotherapy, have been hampered by their side effects. Natural product-derived plants have been used for decades to treat diseases. Compared to conventional therapy, natural product has the potential to be effective against cancer with fewer side effects. This current review discussed the potential of scopoletin, a coumarin present in many edible plants, which elicits anticancer properties through multiple mechanisms, including modulating cell cycle arrest, inducing apoptosis, and regulating multiple signaling pathways. Understanding these mechanisms highlights the potential of scopoletin as a candidate for chemoprevention and chemotherapy.
KEYWORDS: Scopoletin; Coumarin; Anticancer; Signaling pathway; Cell cycle arrest; Apoptosis
1. Introduction
Cancer is a severe global public health concern with increasing morbidity and mortality rates. According to the International Agency for Research on Cancer, the estimated number of new cancer cases worldwide was 19.29 million in 2020, with 9.96 million deaths. In 2040, it is estimated that the number of new cancer cases will be increased by 21%[1], making cancer will soon surpass heart disease as the leading cause of death. Some common treatment strategies for cancer patients, including targeted therapy, chemotherapy, radiotherapy, and surgery[2], have been used to fight against cancer. However, these strategies are limited due to their side effects, lack of specificity, and drug resistance. In addition, they result in poor prognosis and high recurrence rates[3].
To overcome this problem, natural products are continuously developed and used to uncover novel bioactive compounds for anticancer candidates. Natural products have been employed for years to perform disease therapy, including cancer. Natural compounds-derived plants comprise different chemical structures, thus demonstrating a wide range of effects that can be beneficial for complementary or alternative medicine. To date, plant-derived anticancer drugs such as paclitaxel and camptothecin are widely known used for cancer treatment[4]. Natural products are of significant medicinal value in developing anticancer agents, and many available compounds in plants are potentially explored.
Scopoletin is a natural product belonging to the coumarin family that is considered an active ingredient in some plants. It has gained attention for its antitumor properties and great potential as an anticancer by inducing apoptosis and blocking cell proliferation[5-7]. This review summarized the potent anticancer of scopoletin and its underlying mechanisms based on experimental evidence to facilitate potential therapeutic targets of scopoletin in cancer prevention and therapy.
2. Overview of scopoletin
Scopoletin (6-methoxy-7-hydroxycoumarin) is a fluorescent coumarin with a molecular formula of C10H8O4and a molecular weight of 192.16 g/mol. Scopoletin is widely distributed in many medicinal plants within various genera and was reported to have many beneficial effects on human health. Scopoletin has two aromatics rings, one of which is substituted with a hydroxyl group, a methoxy group, and an oxo group (Figure 1). In Arabidopsis, scopoletin is synthesized primarily from the phenylpropanoid pathway via ortho-hydroxylation of cinnamate, p-coumarate, caffeate, and ferulate[8,9]. Reported sources of scopoletin are tabulated in Table 1.
Scopoletin has been extracted from various plants using several extraction methods with different organic solvents. Several extraction methods such as maceration, supercritical fluid extraction, microwave-assisted extraction, reflux, and ultrasonicassisted extraction have been developed to achieve a high yield of scopoletin[10,11]. Novel extraction processes were found to be more efficient for scopoletin extraction, with the advantages of simple, less organic solvent and high yield[12]. Multiple pharmacological actions of scopoletin have been elucidated, including antioxidant[13], antidiabetes[14], antiinflammation[15], and anticancer[5]. Therefore, scopoletin and its derivatives gained the attention of many researchers. This review will describe the underlying mechanisms of scopoletin as an anticancer agent in the following part.
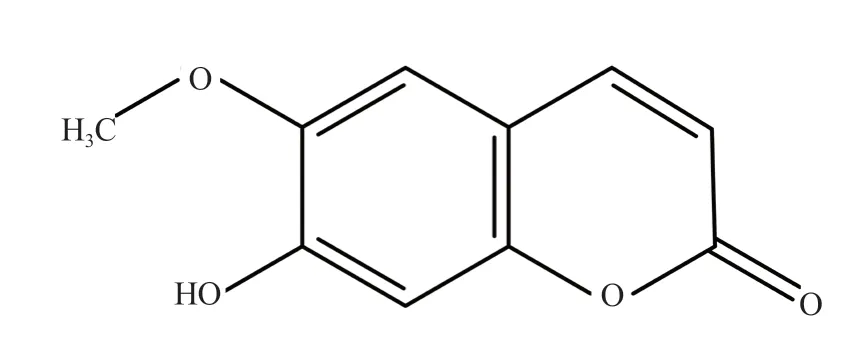
Figure 1. Structure of scopoletin.

Table 1. Sources of scopoletin.
3. Anticancer mechanisms
3.1. Cell cycle arrest
The cell cycle in eukaryotes consists of four discrete phases, termed G0/G1, S, G2, and M phases. G1and G2, the so-called gap phases, are essential in cell proliferation control. G1corresponds to the interval between mitosis and initiation of DNA replication and is followed by S phase for DNA replication. This process is followed by cell growth and protein synthesis in the G2phase, which is further utilized to prepare the M phase for cell mitosis[35]. Protein kinases cyclin dependent-kinases (CDKs) are responsible for modulating the cell cycle of eukaryotes[36]. These protein kinases are activated by cyclins responsible for regulating DNA transcription and replication, mitotic progression, epigenetic regulation, and metabolism[37]. Some cyclin-CDK complexes take place in the cell cycle process, including cyclin C-CDK3 in cell-cycle progression from G0to G1phase and other cyclins (cyclin D-CDK4/6 and B-CDK1) in G1/S and G2/M transitions[38-40]. Consequently, the abnormal activation between these proteins results in cell-cycle dysregulation that leads to malignancies[41].
Scopoletin exposure upregulates cell cycle arrest in cancer cells, including prostate[42], cervical[5], cholangiocarcinoma[7,43], and breast cancer[44]. Scopoletin treatment increased cell cycle arrest in human cervical cancer HeLa and SiHa cells by a different mechanism. Scopoletin resulted in cell accumulation in G2[5] and G0/G1phases in HeLa and SiHa cells, respectively[45]. Similarly, another study by Li et al. also revealed that scopoletin treatment reduced androgen stimulation and caused an accumulation of human prostate cell cancer (LNCaP) and HeLa cells at G2/M checkpoint by decreasing the cyclin D1 expression level[5,42]. In human breast cancer cells, treatment with scopoletin increased cell accumulation in G2/M phase, while in G1and S phases, the percentage of cells markedly decreased[44]. In addition to downregulation of cell cycle regulatory proteins, scopoletin derivative could increase the expression of p21, the downstream effector of p53, and CDK inhibitor. Many reports regarding the potential activity of scopoletin in interfering cell cycle arrest are related to its capability to modulate the expression of cyclin regulatory protein D1, but not B1[42].
3.2. Modulation of PI3K/AKT/mTOR pathway
The phosphatidylinositol-3-kinase (P13K) signaling pathway, which regulates cell growth and differentiation, metabolism, and apoptosis, is the most commonly activated in human cancer[46]. P13K activates serine/threonine-specific protein kinases AKT, its downstream target site, to regulate cell cycle progression. AKT modulates the cell cycle process by activating CDK2 and CDK4 and, at the same time, regulating CDK inhibitor cycle-dependent protein inhibitor P27. P27 protein is highly expressed in normal epithelial tissues such as the lung, breast, ovary, and prostate[47]. Conversely, a loss of P27 protein and dysregulation of the P13K/AKT pathway has been commonly shown in human tumors such as breast, lung, and prostate cancer[48-50].
Moreover, AKT also promotes cell survival by inhibiting apoptosis through various mechanisms, including inactivating deathinducing proteins, repairing cellular DNA damage by activation of transcription factor NF-κB, and inhibiting cytochrome C and apoptotic factors released from the mitochondria[51,52]. AKT activation prompts mTOR, a downstream target site, which then promotes the transcription of ribosomal protein p70 and translation inhibition molecule 4E-BP1. As a primary survival pathway in cancer cells increased, P13K/AKT/mTOR pathway activity correlated with resistance to cancer therapies and tumor progression. Targeting this pathway in cancer is one of therapeutic approaches since many studies reported that inhibiting P13K/AKT/mTORrelated genes and proteins leads to inhibition of human tumors and has been evaluated in many clinical trials at various levels. Therefore, many parties are actively developing inhibitors that target P13K and other critical components in this pathway for cancer treatment[52].
Several studies reported that scopoletin disturbed P13K/AKT/mTOR pathway in cervical[5] and prostate cancer[42]. Scopoletin suppressed the proliferation and migration of HeLa cells. Tian et al.[5] found that scopoletin downregulated the P13K/AKT/mTOR pathway by abrogating the P13K and AKT phosphorylation in HeLa cells. Blocking these pathways induces cell apoptosis and inhibits essential cellular processes in cancer, such as proliferation, migration, and metastasis. Scopoletin treatment also enhanced the expression of apoptotic executioner caspases, such as caspase-3, -8, and -9. A novel scopoletin derivative, SC-Ⅲ3, showed antitumor properties by decreasing mTOR phosphorylation and its downstream effector p70 in human hepatoma HepG2 cells. SC-Ⅲ3 also induced mitochondrial dysfunction leading to autophagy activation, which shows its promising potential as a prospective agent for hepatocellular carcinoma treatment[53].
3.3. Regulation of MAPK pathway
Another pathway responsible for signal transduction regulation is the mitogen-activated protein kinases (MAPKs) pathway that regulates intracellular responses, including cell growth, proliferation, migration, survival, inflammation, and apoptosis. Mammalian MAPKs are classified into p3 kinase, extracellular signal-regulated kinase (ERK), and c-Jun N-terminal kinase (JNK). Phosphorylation of MAPKs was stimulated by intracellular reactive oxygen species (ROS), which is also helpful for the growth and survival of tumor cells. MAPKs are also related to angiogenesis, which is the growth of new blood vessels from the existing ones. This mechanism is vital for many physiological and pathological processes, including embryonic development, bone repair, wound healing, and reproduction[54]. Under physiological conditions, angiogenesis is regulated by a balance between angiogenic modulators. However, uncontrolled angiogenesis results in the formation of diseases including tumor growth and cancer progression[55]. Based on computer modeling, several angiogenic factors, namely ERK1, vascular endothelial growth factor A, and fibroblast growth factor 2, showed high binding energies and ligand affinity to scopoletin[10]. Angiogenesis involves complex signal regulation. However, the antiangiogenic capacities of scopoletin are mainly related to migration inhibition and endothelial cell tube formation, which are correlated with ERK1/2 activation[56].
Scopoletin-mediated reduction in MAPKs phosphorylation has been detected in different kinds of cancer cells, such as the urinary bladder (ECV304 cells)[56], lung (A549 cells)[57], and liver cancer (CCA KKU-100 and HepG2 cells)[58]. Therefore, since MAPKs regulate cellular processes, inactivating them represents a promising strategy, particularly against cancer[59]. Pan et al.[56] reported that scopoletin exposure resulted in ERK1/2 phosphorylation in ECV304 cells, but not p38 and JNK, suggesting that the anti-angiogenic action of scopoletin is likely related to the inhibition of ERK1/2 activation. Scopoletin isolated from Morinda citrifolia (noni) also broadly interfered with the phosphorylation of ERK1/2, JNK, and the α subunit of the eukaryotic translation initiation factor-2 (eIF-2α) in human lung carcinoma A549 cells and cancer-induced mice[60,61]. Kim et al.[62] also revealed that scopoletin downregulated the expressions of matrix metalloproteinase-1 and pro-inflammatory cytokines by inhibiting p38 phosphorylation in human fibroblasts. These findings suggest that scopoletin may protect skin from ultraviolet rays and treat photo-aging, the predominant risk factor causing diseases like cancer[63].
3.4. Apoptosis induction
Apoptosis, called programmed cell death, usually occurs in physiological conditions and is a part of a regulated process. It is well-known to have a significant role in the pathogenesis of diseases since this process leaves clues on how diseases should be treated. Defects in the apoptotic pathways with insufficient apoptosis or downregulation of pro-apoptotic proteins may promote carcinogenesis. Under cancer conditions, cancerous cells lose their apoptotic control and can evade pro-apoptotic mechanisms, thus allowing them to survive longer and metastasize[64]. Cancerassociated defects in apoptosis are responsible for treatment resistance to current traditional cancer therapies, namely chemotherapy, and radiotherapy, requiring a higher dose to eradicate the cancer cells.
Several pathways exist in the programmed cell death mechanism, for instance, the caspase pathway. Caspases are aspartate-specific cysteine proteases responsible for initiating and executing apoptosis[65]. Many studies reported that scopoletin restrained cell growth and increased apoptosis rate dose-dependently in human cervical and promyelocytic leukemia cancer cells[5,6]. Scopoletin exposure significantly increased caspase-3 activity, marked by the upregulation of the active form caspase-3, while procaspase-3 was decreased. Caspase-3 activation further led to the cleavage of poly (ADP-ribose) polymerase (PARP), which was also enhanced in the presence of scopoletin[6]. The induction of PARP cleavage by scopoletin implied the critical function of caspase-3 activation in scopoletin-induced apoptosis.
Shi et al. described that scopoletin and its derivatives initiated apoptosis via a mitochondrial-dependent pathway[66]. The exposure to novel scopoletin derivatives induced depolarization and caused a significant loss of mitochondrial membrane potential in human breast cancer MDA-MB-231 cells[66]. In addition, mitochondrial disruption is also can be affected by oxidative stress. Oxidative stress causes cellular damage by increasing ROS production, opening mitochondrial permeability transition pores to release proapoptotic proteins, cytochrome c, and other caspase-activating proteins[67,68]. The release of activating proteins into the cytosol induces an apoptosis mechanism within the cells. Some proteins are related to the regulation of apoptosis, including Bax and Bcl-2. Scopoletin treatment was reported to result in an increase in Bax and a decline in Bcl-2 expression in many cancer cells, including cervical[5], liver[53], and bile duct cancer cells[43]. Figure 2 depicts the mechanisms of anticancer action of scopoletin.
3.5. Inhibition of multiple drug resistance (MDR)
MDR refers to the resistance or insensitivity to the administrated medicine. In cancer therapy, this condition is a dominant cause of failure in chemotherapy and tumor recurrence. MDR results from defensive mechanisms in cancer, including the genetic mutation of oncogenes that results in drug resistance and the expression of ATPbinding cassette transporters that extrude chemotherapeutic drugs outside the cancer cells[69,70]. Several MDR reversal agents failed to be developed because they suffered resistance in a short time due to severe adverse effects. As a result, there has been significant interest in cancer treatment due to the search for ways in which anticancer drug combinations can act additively or synergistically to increase antitumor activity at lower doses than monotherapy[71,72].
A few works have been conducted to examine the potential use of scopoletin in combination with other chemotherapeutic drugs, for example, cisplatin, to confer synergistic benefits as an anti-cancer. Scopoletin was found to increase the effectiveness of cisplatininduced apoptosis in cholangiocarcinoma[43]. Cisplatin is a wellknown therapeutic drug that treats numerous cancers, including head and neck, ovarian, and testicular cancer. Cisplatin inhibits DNA replication, interferes with DNA repair mechanism, and causes DNA damage, thus leading to DNA apoptosis[73,74]. The cell cycle arrest induced by cisplatin was improved when combined with scopoletin, resulting in a 2-fold increase of apoptosis in cholangiocarcinoma cells[43]. This combination of scopoletin and cisplatin could be beneficial in treating cholangiocarcinoma.
4. Preclinical studies of scopoletin
Although scopoletin exhibits marked antitumor and anticancer activity in in vitro studies, few preclinical and clinical studies are reported. Scopoletin has been shown to exhibit an anticancer effect on a number of cancer cells. Table 2 tabulates the preclinical studies on the dosage, efficacy, and potential molecular mechanisms of scopoletin in the treatment of different cancer cells.

Figure 2. Schematic summary of anticancer mechanisms of scopoletin.
5. Pharmacokinetics, bioavailability, and potential drug delivery of scopoletin
Oral administration is the most acceptable and convenient way of drug administration. However, given the pharmacokinetic and biopharmaceutical studies conducted in humans, scopoletin is lipophilic, poorly water-soluble, poorly absorbed, and rapidly metabolized[79]. As a result, to improve its bioavailability, it may be necessary to boost its concentration by using a co-solvent or a carrier system, modifying the structure, or adding new functional groups. Several efforts have yielded results in the development of efficient drug carriers to deliver scopoletin to the target site, including the polylactic-co-glycolic acid-nanoparticle, loaded with scopoletin[80], gold nanoparticle loaded with scopoletin[81] and polymeric micelles containing scopoletin[82]. These drug delivery methods have successfully improved the solubility and absorption of scopoletin. Scopoletin was encapsulated with polylactic-coglycolic acid and a small-sized nano-scopoletin was yielded with more than 85% entrapment efficiency. This nano-scopoletin resulted in enhanced cellular entry and greater bioavailability of scopoletin and demonstrated a faster cellular uptake than its un-encapsulated counterpart[80]. Oral administration of scopoletin encapsulated in polymer-based micelles also effectively increased the bioavailability of scopoletin in the liver. Polymeric micelles have been applicable to ameliorate the water solubility and oral bioavailability of poorly soluble drugs[83,84] and are promising for improving the oral bioavailability of inadequately soluble compounds, for instance, scopoletin, to achieve better therapeutic effects.
6. Toxicity and adverse effects
Despite the wide range of pharmacological benefits of coumarin, there are few reports about adverse effects from exposure to high doses of the coumarin group, for instance, scopoletin. The toxicity of coumarins was assessed in preclinical studies using animal models. There are no signs of toxicity observed in chronic and subchronic studies on B6C3F1 and CD-1 mice which were treated with 19-300 mg/kg BW and 300-3 000 ppm coumarins, respectively[85,86]. However, signs of liver toxicity were discovered both in chronic and subchronic studies. Oral administration of 50-500 mg/kg BW coumarins for 13 weeks caused liver toxicity in Sprague-Dawley rats[85]. In addition, the same type of rats showed signs of anemia after being administrated with 444-5 000 ppm coumarins for 2 years[87].
7. Conclusion and future perspective
Natural products have received much attention in recent decades as potential sources of new anticancer drugs because they possess different responses in tumor cells and show more practical effects than available chemotherapy drugs. The antitumor action of scopoletin is primarily due to the induction of multiple signal transduction pathways, including cell proliferation inhibition, cell cycle arrest, and apoptosis induction. Since humans widely consume scopoletin-containing foods, further studies with multiple doses of scopoletin in the animal model are required to fully understand its potential and mechanism as an anticancer agent in humans.

Table 2. Antitumor effects of scopoletin in preclinical studies.
Conflict of interest statement
The authors have no conflict of interest to declare.
Funding
The authors received no extramural funding for the study.
Authors’ contributions
Conceptualization: RMD and LDA; literature research: LM, RMD, and LDA; visualization: LM and LDA; writing – original draft preparation: LM, RMD, and LDA; writing – review and editing: ANT, AWS, and LDA. All authors have read and agreed to the published version of the manuscript.
 Asian Pacific Journal of Tropical Biomedicine2023年1期
Asian Pacific Journal of Tropical Biomedicine2023年1期
- Asian Pacific Journal of Tropical Biomedicine的其它文章
- Information for Authors Asian Pacific Journal of Tropical Biomedcine
- Molluscicidal effect of green synthesized silver nanoparticles using Azadirachta indica on Biomphalaria alexandrina snails and Schistosoma mansoni cercariae
- Anti-leishmanial, immunomodulatory and anti-oxidative activity of quercetin against cutaneous leishmaniasis caused by Leishmania major
- L-carvone attenuates myocardial injury and dyslipidemia in rats with isoproterenolinduced cardiac hypertrophy
- Quercetin modulates ovarian autophagy-related molecules and stereological parameters in a rat model of PCOS