
CRISPR/Cas基因编辑技术与植物病毒研究进展
2023-01-19 11:56徐子妍李浩周焕斌周雪平
浙江大学学报(农业与生命科学版) 2022年6期
徐子妍,李浩,周焕斌,周雪平*
(1.浙江大学农业与生物技术学院生物技术研究所,杭州 310058;2.中国农业科学院植物保护研究所,北京 100193)
植物易受到真菌、细菌、病毒、线虫、寄生性植物的侵染胁迫。全球每年因各种病原物和害虫危害导致水稻(Oryza sativa)、玉米(Zea mays)、小麦(Triticum aestivum)、马铃薯(Solanum tuberosum)、大豆(Glycine max)等主要粮食作物产量损失20%~40%,而近半数的经济损失由植物病毒病造成,每年高达300亿美元[1]。植物病毒病害因难以防治的特性而素有“植物癌症”之称,病害发生时会严重影响农作物的正常生长发育,使感病植株出现矮化、萎缩、花叶、缺绿、斑驳、坏死、丛簇等症状,严重时会导致寄主死亡,影响农作物产量与品质,威胁农业生产及粮食安全。多年来,由于缺乏抗性品种及有效的化学农药,植物病毒病害多以预防为主,即采用培育无毒种子种苗、清除病株、切断昆虫介体传播途径等措施,因此开发广谱、高效的抗病毒品种是当务之急。
规律成簇的间隔短回文重复序列及其相关蛋白[clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein (Cas),CRISPR/Cas]系统是古细菌或细菌抵抗病毒以及其他外来遗传元件的免疫系统。近年来,基于CRISPR/Cas的基因编辑技术快速发展,已经被广泛应用于目的基因敲除、转录调控、碱基编辑、DNA片段插入和替换等多个研究领域[2]。这一革命性的技术可应用于农作物粮食增产、品质改良、抗性提升等多个育种方向,尤其可以提升农作物抵御病原物侵染的能力,减少病原物对农作物产量和品质的危害。
植物病毒基因组根据构成的不同,可分为DNA病毒和RNA病毒。得益于CRISPR/Cas基因编辑技术的不断发展,目前已有多种Cas 蛋白可分别靶向DNA病毒序列和RNA病毒序列,通过切割病毒基因组的关键序列,抑制其侵染、复制、转录等多个过程,从而提高携带CRISPR/Cas 基因编辑材料寄主对病毒的抵抗力。同时,CRISPR/Cas基因编辑技术还可以靶向寄主因子,通过敲除感病基因来抑制病毒的侵染,亦可提高寄主抗性[3]。此外,植物病毒基因组因相对简单、方便改装、易于侵染,也被应用于传递CRISPR/Cas,完成对寄主靶基因的编辑[4],该策略可绕开植物组培转化操作,进一步节省人力、物力。最新研究成果显示,多种病毒已经被成功用于递送核酸酶等大分子物质进入寄主,缩短了基因编辑周期[5-6]。
1 CRISPR/Cas 基因编辑技术
CRISPR/Cas 是一类来源于细菌或古细菌中的适应性免疫防御系统。由于其具有精准靶向基因序列的特征,因此被开发为高效的基因编辑工具。与前两代的锌指核酸酶(zinc finger nucleases,ZFNs)基因编辑技术和类转录激活因子效应物核酸酶(transcription activator-like effector nucleases,TALENs)基因编辑技术相比,CRISPR/Cas 基因编辑技术工作原理更为简单,主要通过引导RNA(guide RNA,gRNA)引导Cas 蛋白至特异性位点切割DNA 序列,并通过内源的同源定向修复(homology-directed repair,HDR)和非同源末端连接(non-homologous end joining, NHEJ)修复,从而产生碱基的插入或缺失,具有操作门槛低、编辑效率高、成果转化快等优点,目前已成为农作物育种、基因功能研究的重要工具[7]。
根据Cas基因主要元件序列的不同,CRISPR/Cas基因编辑技术目前被分为2大类[分类1(Class 1)和分类2(Class 2)],其中Class 1系统(包括Ⅰ、Ⅲ和Ⅳ型)的基因编辑利用由多个效应蛋白组成的复合物行使功能,而Class 2系统(包括Ⅱ、Ⅴ和Ⅵ型)仅需一个效应蛋白[8]。被广泛应用的CRISPR/Cas9 属于Class 2系统中的Ⅱ型,仅由gRNA和Cas9蛋白2部分组成。其编辑载体的构建步骤非常简单,只需根据靶向序列设计特异性的、长度为19~20 bp的单向导RNA(single guide RNA,sgRNA),就可以引导Cas9蛋白到特异性位点实现切割。由于Cas蛋白结合靶序列需要特定的前间区序列邻近基序(protospacer adjacent motif,PAM),而CRIPSR/Cas9蛋白只能识别NGG的PAM位点,无法识别其他位点,这极大地限制了其应用范围。因此,通过开发多种CRIPSR/Cas9的变体及同源蛋白来拓宽其在动植物中的编辑范围很有必要。如Cas9 变体CRIPSR/Cas9-NG、SpCas9-EQR、SpCas9-VQR、SpCas9-VEQR、SpCas9/SpG、SpCas9/SpRY 等能够识别NGN、NGAG、NGA、NGCG、NGN、NNN的PAM位点[9];同时,Cas9的同源蛋白SaCas9(金黄色葡萄球菌,Staphylococcus aureus)、ScCas9(犬 链 球 菌,Streptococcus canis)、St1Cas9(嗜热链球菌,Streptococcus thermophiles)、BlatCas9(侧孢短芽孢杆菌,Brevibacillus laterosporus)等能够有效识别NNG、NNGRRT、NNAGAW、NNNCND的PAM位点[9]。
此外,以CRISPR/Cas12 为代表的新型基因编辑技术也应运而生。CRISPR/Cas12 属于Class 2 系统中的Ⅴ型[10],仅由源于CRISPR 的RNA(CRISPRderived RNA,crRNA)和Cas蛋白2部分组成并发挥作用。与Cas9相比,Cas12蛋白具有分子量更小、分子之间连接更紧凑、脱靶效率更低并且能够识别富含T的PAM等优点。这些CRISPR系统大大拓展了科研工作者的基因编辑工具箱,丰富了其在植物基因编辑中的应用。
以上所述Cas 蛋白均靶向DNA 序列,无法对RNA 序列进行编辑。2016 年,ABUDAYYEH 等首次报道了一种新的Cas蛋白,命名为Cas13a,它属于Class 2系统中的Ⅵ型蛋白[11]。与Cas12类似,Cas13a同样由crRNA和Cas蛋白2部分组成并发挥编辑功能[12]。其工作原理与其他编辑系统相似,通过crRNA 将Cas13a 引导至RNA 序列特定位点进行编辑。这套系统实现了对RNA序列的编辑,在更好地研究生物体内RNA的各种调控作用,尤其是在植物寄主中RNA 病毒的防控以及农作物育种等研究方向上具有里程碑式意义。
除了天然的Cas 蛋白核酸酶,经过改造的Cas9蛋白(dCas9和nCas9)也被广泛用于单碱基编辑、引导编辑、转录调控等。CRISPR/Cas介导的碱基编辑技术可在动植物体内实现高效的单碱基替换,它由dCas9和胞嘧啶脱氨酶或者腺嘌呤脱氨酶组成,从而能实现从C→T 或者从A→G 的替换[13]。与HDR 相比,碱基编辑技术不需要DNA双链断裂以及供体模板即可实现高效的单碱基替换,对编辑对象尤其是在提高农作物抗性的同时对其农艺性状影响更小,因此,在未来的作物育种研究中单碱基编辑的可操作性更强。此外,利用dCas9携带甲基化、去甲基化等相关基因核心元件,在不切割基因序列的情况下,可以将该元件靶向至定点区域,实现精确的甲基化修饰等过程,调控基因开关,实现定向调控。
目前,CRISPR/Cas基因编辑技术已被应用于包括水稻、番茄(Solanum lycopersicum)、玉米、小麦等多种农作物的性状改良、抗性提升等研究中,给未来农作物基因功能研究以及作物育种带来了更多的可能性,尤其是单碱基编辑技术的开发,将为多种农艺性状精确矫正、作物增产、病虫害防治带来曙光。
2 基因编辑技术在植物抗病毒研究中的应用
目前,CRISPR/Cas基因编辑技术不仅被应用于植物基因组编辑工作,而且已被广泛用于植物抗病毒工程研究中,是除传统抗病毒策略外最具发掘潜力和技术优势的抗病毒手段,对于后续抗病毒育种研究具有重要的指导意义。根据编辑对象的不同,目前利用CRISPR/Cas 基因编辑技术防治病毒的主要策略可以分为2种:一是携带CRISPR/Cas的寄主直接靶向病毒基因序列,通过破坏病毒功能性序列来抵抗病毒侵染;二是病毒若要成功侵染植物并进行复制、增殖,则需要利用多种寄主内源基因协助其完成侵染、复制、转录、组装等整个过程,因此可通过靶向此类寄主植物的内源基因,以阻碍病毒侵染、复制、增殖等多个环节,从而达到抵抗病毒、降低病毒积累量的目的(图1和表1)。
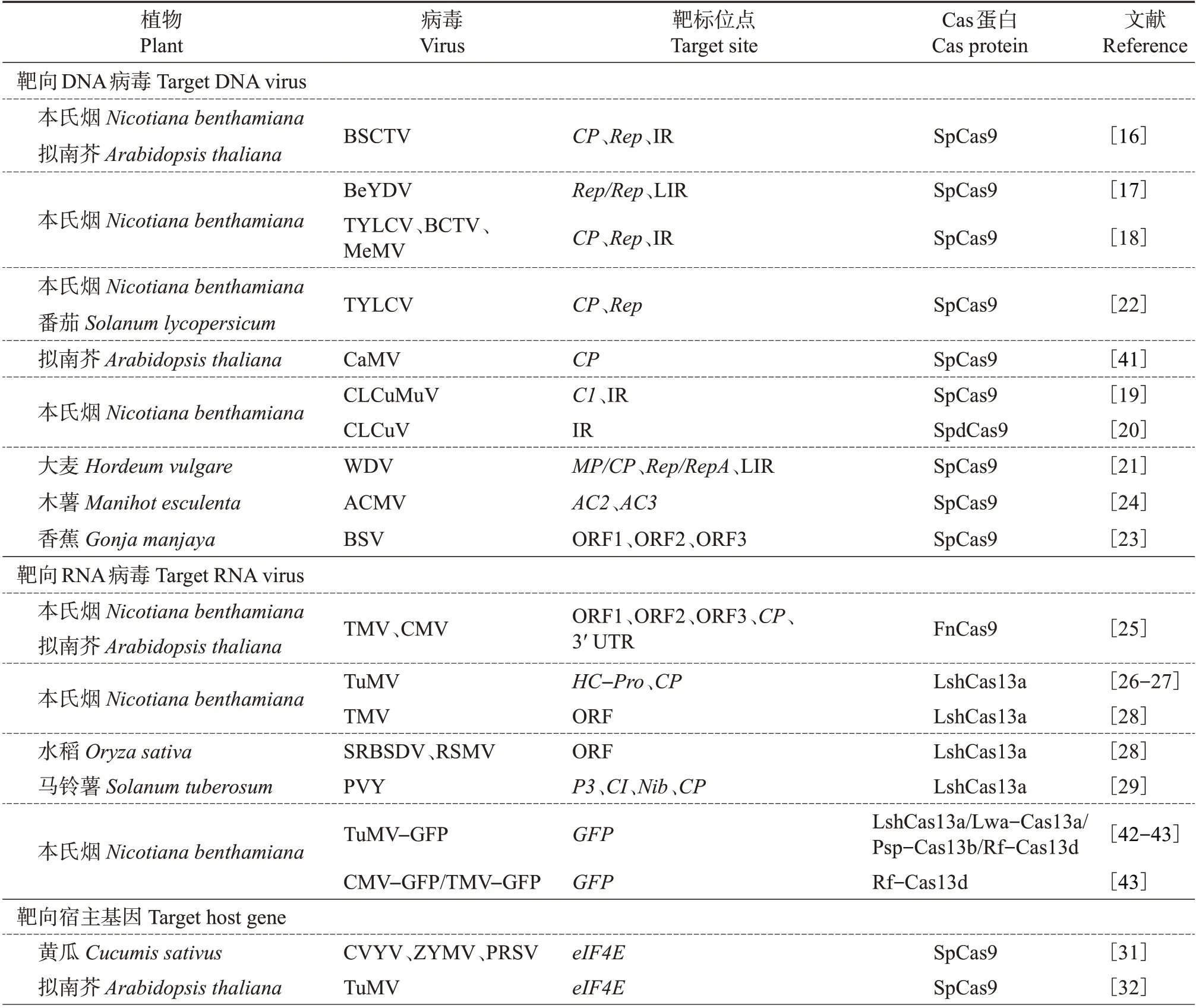
表1 CRISPR/Cas基因编辑技术在植物抗病毒中的应用Table 1 Application of CRISPR/Cas gene editing technology on plant resistance to viruses

表1(续) Continuation of Table 1
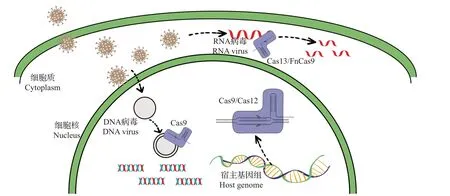
图1 CRISPR/Cas基因编辑技术应用下的抗病毒策略Fig.1 Antiviral strategy under the application of CRISPR/Cas gene editing technology
2.1 靶向病毒基因组
2.1.1 DNA 病毒
在植物中,根据基因组核酸的不同,DNA 病毒可分为双链DNA 病毒和单链DNA 病毒。其中,双链DNA 病毒比较少见,主要为花椰菜花叶病毒科(Caulimoviridae)病毒,该病毒科寄主范围较窄,主要侵染花椰菜(Brassica oleracea)、大豆、水稻等。该病毒科代表种花椰菜花叶病毒属于花椰菜花叶病毒属(Caulimovirus),主要由蚜虫以半持久性方式传播,也能以机械方式传播接种[14]。单链DNA病毒以双生病毒科(Geminiviridae)研究最为广泛,该病毒科在全世界范围内广泛存在,能够侵染禾本科(Gramineae)、茄科(Solanaceae)、葫芦科(Cucurbitaceae)和豆科(Leguminosae)等多种重要农作物,导致农作物产量和品质严重下降,对我国乃至世界的农业发展及粮食安全带来重大打击[15]。双生病毒以滚环复制的方式进行增殖,即以单链DNA 为模板,形成闭合环状双链DNA 中间体,并以此进行滚环复制扩增子代DNA来进行增殖,多通过粉虱科(Aleyrodidae)、蚜总科(Aphidoidea)、叶蝉科(Cicadellidae)等昆虫介体进行传播。为了抵抗双生病毒对植物引起的病害,JI等针对甜菜严重曲顶病毒(beet severe curly top virus,BSCTV)基因组的编码区和非编码区设计了43个靶标位点,通过瞬时表达实验证明不同靶标位点的病毒积累量都不同程度地减少,进一步发现稳定表达sgRNA/Cas9 转基因的拟南芥(Arabidopsis thaliana)和本氏烟(Nicotiana benthamiana)对BSCTV 表现出高度抗性[16]。BALTES等在烟草中针对菜豆黄矮病毒(bean yellow dwarf virus,BeYDV)基因组的复制起始蛋白(replication initiator protein, Rep)多个结合位点和滚环复制所必需的3 个Rep 基序(基序Ⅰ、Ⅱ和Ⅲ)设计了11 个靶标位点,使得本氏烟中的病毒积累量降低了87%,从而赋予了植物对BeYDV的抗性[17]。ALI等针对病毒的衣壳蛋白(coat protein, CP)、Rep 和基因间隔区(intergenic region,IR)设计了sgRNA,结果显示:靶向番茄黄化曲叶病毒(tomato yellow leaf curl virus,TYLCV)的IR区域能够显著降低病毒积累量,减轻病害症状;双生病毒的IR区域存在保守的TAATATAC序列,通过设计针对该保守序列的IR-sgRNA,能够同时靶向TYLCV、甜菜曲顶病毒(beet curly top virus,BCTV)和鱼黄草花叶病毒(merremia mosaic virus, MeMV)基因组,从而获得抵御多种病毒侵染的广谱抗性[18]。使用类似的方法,稳定表达Cas9蛋白靶向木尔坦棉花曲叶病毒(cotton leaf curl Multan virus, CLCuMuV)编码的C1蛋白和IR区域的双gRNA 转基因烟草被赋予对该病毒侵染的抵抗能力[19]。KHAN等获得了表达dCas9-sgRNA的转基因本氏烟材料,验证结果表明,在没有切割基因组的情况下该材料能够有效抑制棉花曲叶病毒(cotton leaf curl virus,CLCuV)的复制[20]。此种策略同时在大麦(Hordeum vulgare)、番茄、香蕉(Gonja manjaya)的抗病毒工程研究中得到了检验,在不同寄主中分别针对小麦矮缩病毒(wheat dwarf virus, WDV)[21]、TYLCV[22]和香蕉线条病毒(banana streak virus,BSV)[23]等病毒基因组设计特异性sgRNA进行定点靶向,达到了阻碍病毒复制、减少病毒积累量的目的。
有研究表明,CRISPR/Cas9 基因编辑技术对双生病毒的编码区和非编码区表现出不同的编辑效率,且不同双生病毒编码区的同源定向修复非常高效,因此通过靶向病毒的编码区,可能会引起病毒重组形成新的病毒变体,从而逃避CRISPR/Cas9 的识别与编辑。MEHTA 等[24]通过靶向非洲木薯花叶病毒(African cassava mosaic virus,ACMV)的AC2 和AC3蛋白编码序列获得了Cas9转基因木薯材料,与对照组相比,实验组并没有表现出明显的抗性,进一步测序发现,有33%~48%的被编辑病毒在靶标识别位点进化出1 个保守的单核苷酸突变,并通过这种突变逃避sgRNA的识别,避免被CRISPR/Cas9切割。这种病毒通过突变逃避编辑的现象需引起科研工作者在抗病毒工程研究中对基因编辑技术的重视。
2.1.2 RNA 病毒
根据基因组的不同,RNA病毒可分为单链RNA(single-stranded RNA, ssRNA)和双链RNA(doublestranded RNA, dsRNA)病毒。其中,ssRNA 病毒进一步可以分为正义ssRNA(+ssRNA)病毒和负义ssRNA(-ssRNA)病毒[14]。最近,有研究发现Cas9蛋白突变体以及Cas13蛋白能够切割RNA链,据此利用CRISPR/FnCas9 针对黄瓜花叶病毒(cucumber mosaic virus,CMV)和烟草花叶病毒(tobacco mosaic virus,TMV)的基因组设计了sgRNA,并获得了稳定遗传的烟草和拟南芥,其后代病毒感染症状显著减弱,病毒积累量显著减少[25]。此外,利用CRISPR/Cas13a对融合绿色荧光蛋白(green fluorescent protein,GFP)作为荧光标签的芜菁花叶病毒(turnip mosaic virus,TuMV)基因组进行靶向发现,编辑病毒基因组的不同区域均有效降低了GFP表达量,同时减少了系统叶片中TuMV-GFP RNA基因组的积累,干扰了TuMV 病毒的复制和传播[26-27]。ZHANG 等[28]利用CRISPR/Cas13a(LshCas13a)在双子叶植物烟草中靶向TMV RNA基因组的同时,赋予单子叶植物水稻对南方水稻黑条矮缩病毒(southern rice black-streaked dwarf virus, SRBSDV)和水稻条纹花叶病毒(rice stripe mosaic virus,RSMV)的抗性。而利用CRISPR/Cas13a 编辑马铃薯Y 病毒(potato virus Y, PVY)基因组的保守编码区,其转基因材料表现出对多种PVY变体的广谱抗性[29]。
2.2 靶向宿主植物内源基因
植物病毒除编码复制酶外,并不编码其他复制相关蛋白,因此需要依赖于宿主植物复制相关因子完成自身复制。此外,病毒的转录、翻译及移动也需要寄主因子进行协作,而这些基因大多为植物感病基因,因此可以通过编辑该类基因抑制病毒复制等环节,进一步减少病毒积累量,降低致病性。2002年,研究人员在拟南芥中首次发现了由真核翻译起始因子4E(eukaryotie translation initiation factor 4E,eIF4E)介导的隐形抗病毒基因,eIF(iso)4E突变体表现出对烟草蚀刻病毒(tobacco etch virus,TEV)的高抗性[30]。辣椒(Capsicum annuum)、野番茄(Solanum habrochaites)、莴苣(Lactuca sativa)、甜瓜(Cucumis melo)、水稻和大麦中的eIF4E也可获得对马铃薯病毒的抗性。此外,eIF4E还介导寄主对多种病毒的抗性,例如黄瓜花叶病毒、芜菁皱缩病毒(turnip crinkle virus,TCV)、甜瓜坏死斑病毒(melon necrotic spot virus, MNSV)、大麦黄花叶病毒(barley yellow mosaic virus, BYMV)、大麦轻度花叶病毒(barley mild mosaic virus,BaMMV)和水稻黄斑驳病毒(rice yellow mottle virus,RYMV)。
2016 年,CHANDRASEKARAN 等[31]首次利用CRISPR/Cas9 对黄瓜(Cucumis sativus)的eIF4E基因进行敲除,获得的T3代非转基因纯合突变体对黄瓜脉黄化病毒(cucumber vein yellow virus,CVYV)、小西葫芦黄花叶病毒(zucchini yellow mosaic virus,ZYMV)和番木瓜环斑病毒(papaya ring spot virus,PRSV)具有广谱抗性。此外,利用CRISPR/Cas9 对拟南芥中的eIF4E进行突变,其突变体对TuMV具有抗性[32];水稻中的eIF(iso)4E进行突变后可抵御水稻东格鲁球状病毒(rice tungro spherical virus,RTSV)侵染[33]。GOMEZ等利用CRISPR/Cas9同时对木薯中的eIF4E异构体nCBP-1和nCBP-2进行敲除,此举措增强了植株对木薯褐色线条病毒(cassava brown streak virus,CBSV)的抗病能力[34]。
eIF4E基因在植物的翻译起始过程中发挥着重要作用,且eIF4E家族存在高度功能冗余,马铃薯病毒能够选择性地利用eIF4E基因或eIF(iso)4E基因或两者兼用来完成增殖过程,因此敲除其中一个基因不足以让植物产生高度抗性,只有同时敲除多个eIF4E基因才能赋予植物最强的抗病毒能力,但同时也会显著影响植物的正常生长发育[35]。挖掘天然抗性等位基因的关键位点,在赋予植物广谱抗病毒能力的同时不影响植物的生长发育尤为重要,这也对植物抗病毒育种提出了更高要求。2019 年,BASTET等利用CRISPR/Cas9介导的碱基编辑技术实现了将拟南芥中eIF4E中的第176位天冬氨酸转变为赖氨酸,从而获得了对三叶草黄脉病毒(clover yellow vein virus, ClYVV)具有抗性且正常生长发育的遗传材料,弥补了无法对eIF4E多基因进行编辑的空缺[36]。
卡哈尔体(Cajal bodies, CBs)是真核细胞中独特的亚核结构,通常与核仁功能相关。CBs在RNA代谢、转录、剪接,核糖体生物合成以及端粒维持的核糖核蛋白(ribonucleoprotein,RNP)形成中发挥着重要作用,同时CBs 还参与细胞周期、发育和应激反应的调节。而Colin 作为卡哈尔体的特征蛋白,是其形成和维持功能活性所必需的结构支架蛋白,并且Colin 蛋白能够促进PVY 病毒的复制,增强其致病性。MAKHOTENKO等通过基因枪将CRISPR/Cas9-sgRNA 递送到马铃薯的顶端分生组织以对Colin基因进行编辑,其突变体后代对PVY 的抗病性有所提高[37]。
由大豆花叶病毒(soybean mosaic virus, SMV)引起的大豆花叶病是限制全球大豆产业健康发展的重要病害之一。大豆异黄酮合酶(Glycine maxisoflavones synthase, GmIFS)的多态性可引起大豆异黄酮积累量的增加,增强寄主对SMV 抗性[38]。ZHANG 等利用CRISPR/Cas9 同时对大豆中的GmF3H1、GmF3H2和GmFNSII-1基因进行敲除,提高了大豆异黄酮的积累量,3 靶点突变体大豆对SMV抗性提升[39]。
番茄褐色皱果病毒(tomato brown rugose fruit virus,ToBRFV)是烟草花叶病毒属中的一种新兴病毒,2014 年在中东地区被首次报道,目前已在全球范围内广泛传播。鉴于其对番茄种植产业造成的巨大危害,2021 年被我国农业农村部列入《中华人民共和国进境植物检疫性有害生物名录》(http://www.zys.moa.gov.cn/flfg/201904/t20190428_6245344.htm),因此,创制出抗ToBRFV 的新型种质资源迫在眉睫。拟南芥中编码的TOBAMOVIRUS MULTIPLICATION1(TOM1)基因是烟草花叶病毒属病毒复制和增殖所必需的,其突变后能显著抑制TMV 的积累。ISHIKAWA 等[40]通过CRISPR/Cas9基因编辑技术对番茄中TOM1的同源基因进行敲除,赋予了番茄对ToBRFV的抗性。
3 植物病毒递送CRISPR 元件的建立与优化
随着对植物病毒基因组功能以及植物与病毒互作机制的深入研究,多种植物病毒已被成功开发为生物技术研究工具。寄主植物本身对病毒具有遗传免疫机制,根据其转录后基因沉默(posttranscriptional gene silencing, PTGS)的机制可以令病毒携带寄主的基因片段,在病毒侵染激活寄主自身免疫系统降解病毒RNA 的同时也可以降解目的基因的微RNA(microRNA),达到识别并抑制该基因表达的目的。烟草脆裂病毒(tobacco rattle virus,TRV)能够侵染50 科400 多种寄主,介导本氏烟、番茄等多种寄主植物发生病毒诱导的基因沉默(virusinduced gene silencing, VIGS)。目前,多种植物病毒已被证实可作为RNA沉默载体,可用于多种农作物基因的功能研究。植物病毒也可用于表达外源蛋白,从而控制农作物农艺性状或者进行疫苗生产。随着基因编辑技术的迅速发展,病毒可通过递送CRISPR元件进入寄主体内实现对寄主特定基因的编辑。
3.1 DNA 病 毒 递 送CRISPR 元 件
双生病毒作为DNA病毒的重要成员,具有2.5~3.0 kb 的单链环状DNA 基因组。通过双生病毒传递CRISPR/Cas 系统具有多种优势:首先,双生病毒寄主范围广,能够侵染多种植物,同一病毒载体能够应用于多种寄主的基因编辑(表2);其次,双生病毒的复制相关蛋白(Rep)可在宿主细胞内启动复制,加快病毒复制速度,提高基因编辑效率;再者,双生病毒可在宿主体内通过高效复制产生大量序列特异性核酸酶(sequence-specific nucleases,SSNs)和gRNA,大大提高靶向效率;最后,双生病毒在宿主细胞内依赖同源重组进行复制,因此可通过外源增加SSNs和gRNA提高同源定向修复效率。
双生病毒基因组小,易于操作,但这也限制了双生病毒用作载体的载货能力。通过替换双生病毒CP蛋白可使双生病毒能够携带800~1 000 bp的外源蛋白序列,同时不影响其运动和复制能力。然而,CRISPR/Cas 蛋白多为4 000 bp 左右的外源蛋白,双生病毒无法携带此类大分子蛋白。为了增强双生病毒的载货能力,需同时去除其运动蛋白和衣壳蛋白,形成非侵染性复制子,并通过农杆菌介导的遗传转化递送至植物细胞中,这大大提高了双生病毒在植物中的HDR效率。
BALTES 等首次利用BeYDV 递送ZFN 和修复模板进入烟草细胞,实现了目的基因的有效编辑,并进一步利用BeYDV 复制子递送TALEN 和CRISPR/Cas9 元件验证了双生病毒复制子具有足够的承载能力,提高了其在烟草中的编辑效率[44]。ERMÁK等利用BeYDV复制子在调节番茄花青素生物合成的基因上游插入一个强启动子,使得番茄组织中花青素的含量显著提高,与传统的DNA递送方法相比,使用双生病毒复制子的方法可使编辑效率提高近10倍[45]。VU等基于BeYDV复制子创建了依赖于CRISPR/LbCpf1系统的同源重组载体表达系统,效率较Cas9 蛋白介导的同源系统提高了3~4倍[46]。GIL-HUMANES等开发了基于WDV复制子精准靶向谷类作物基因的编辑系统,由该双生病毒复制子介导的报告基因在小麦中的表达量增加了110倍,且与非病毒递送方法相比,利用其携带CRISPR/Cas9 核酸酶和修复模板让小麦中的基因编辑效率提高了12倍,并可在小麦的同一细胞中实现多重基因靶向,虽然效率仅有1%[47]。WANG 等使用WDV复制子系统在水稻中实现了由CRISPR/Cas9介导的基因敲入,其工作效率进一步提高至19.4%[48]。双生病毒复制子介导的基因编辑不仅能够在烟草、番茄、小麦和水稻中实现有效编辑,而且同样适用于马铃薯基因组研究。BUTLER 等在2015 年和2016 年发表的2项研究结果证实了双生病毒复制子介导的基因编辑的可行性并展望了其广阔的应用前景[49-50]。
除用双生病毒复制子系统来传递表达CRISPR/Cas9 外,YIN 等首次利用双生病毒载体开发了病毒诱导型基因组编辑系统(virus-induced genome editing system,VIGE),通过双生病毒载体递送gRNA 至转基因Cas9的本氏烟中,实现了对NbPDS3和NbIspH内源基因的有效打靶[51]。
3.2 RNA 病 毒 递 送CRISPR 元 件
除了DNA 病毒,许多RNA 病毒也被用作植物中递送元件的载体(表2)。TRV 具有2 条正向单链RNA:RNA1和RNA2。RNA1对病毒复制和运动至关重要,而RNA2则编码CP、2b、2c蛋白。通常情况下,通过对RNA2中的2b和2c蛋白进行删除从而实现外源蛋白表达。利用TRV 可以将ZFN 元件递送至植物的各种组织和细胞中,并引起有效的基因编辑,从而避免植物遗传转化过程,缩短育种时间[52]。2015 年,TRV 首次被应用于CRISPR/Cas 基因编辑载体的递送:ALI等通过农杆菌侵染的方法,将带有八氢番茄红素去饱和酶(phytoene desaturase,PDS)基因的sgRNA TRV载体递送至过表达Cas9的本氏烟转基因品系的叶片中,引起了PDS 的突变,并且TRV能够侵染分生组织,由此可知其当代种子稳定携带了该遗传修饰,并可遗传给后代[53]。此TRV递送系统可用于抗病毒研究,通过携带多个靶向TYLCV 基因组的sgRNA 可获得抗TYLCV 的转基因本氏烟[54]。TRV 同样可以携带crRNA,将其传递到过表达CRISPR/pCas13a的本氏烟中,使本氏烟能有效抵抗TuMV 的侵染[26]。通过在过表达Cas9 的本氏烟中比较TRV 和豌豆早褐病毒(pea earlybrowning virus, PEBV)载体递送sgRNA 效率时发现:PEBV 介导的基因编辑具有更高的编辑效率(57%~63%);同时,PEBV同样可以感染分生组织,这就意味着不需要传统的组织培养即可获得可遗传的靶向突变[55]。
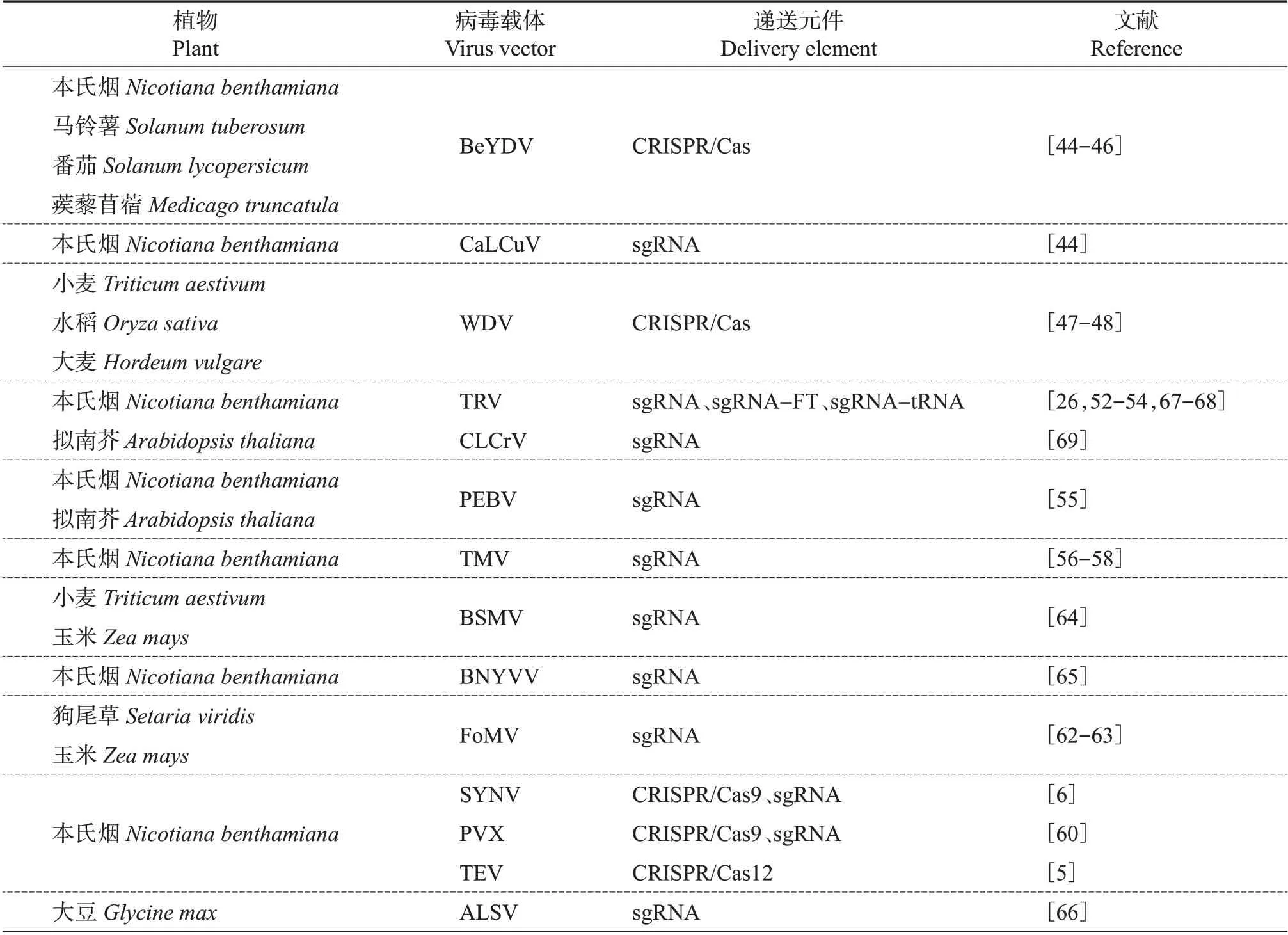
表2 病毒诱导的基因编辑在植物中的应用Table 2 Application of virus-induced gene editing(VIGE)on plants
TMV 也被开发为递送sgRNA 的载体,通过TMV 载体递送高浓度sgRNA 编辑本氏烟内源基因,其编辑效率高达70%;TMV 载体也能够进行多重基因组编辑,以及同一载体同时递送sgRNA和过表达某一基因,做到基因敲除与基因过表达同时进行[56-57]。为进一步优化TMV诱导的基因编辑系统,CHIONG 等利用2 个TMV 载体分别递送Cas9 和sgRNA 时在烟草中引起了插入缺失,而单个TMV载体虽然可同时递送Cas9 和sgRNA,但与前者相比,其在烟草中的编辑效率显著较低,仅为7%[58]。
马铃薯X 病毒属(Potexvirus)的病毒也可作为sgRNA 递送载体,马铃薯X 病毒(potato X virus,PVX)是正单链RNA 线性病毒,长度为6 345 bp[59],能够同时携带Cas9 和sgRNA 并递送至本氏烟中,造成有效突变。PVX 也可递送胞嘧啶碱基编辑器(cytosine base editor,CBE)载体,该系统在NbTOM1处引起了C→T 的碱基替换,证实PVX 载体可与CBE兼容[60]。同属于马铃薯X病毒属的狗尾草花叶病毒(foxtail mosaic virus,FoMV)也已被开发为VIGS载体和基因表达载体[61]。利用FoMV 载体携带sgRNA,可在玉米、狗尾草(Setaria viridis)和本氏烟中实现基因编辑,但使用FoMV系统靶向玉米耐盐基因ZmHKT1的编辑效率仅为3%~6%[62-63]。当FoMV与甘蔗花叶病毒(sugarcane mosaic virus,ScMV)共同接种时,表现出对狗尾草SvCA2基因更高的编辑效率,其中接种叶为45%,系统叶为60%,但这些编辑是不可遗传的[63]。URANGA 等提出了使用2 种兼容的病毒载体共同表达Cas 蛋白和sgRNA,通过将表达LbCas12a 的TEV 载体和表达sgRNA 的PVX载体农杆菌共同浸润烟草,获得了无转移DNA(transferred DNA,T-DNA)插入的基因编辑植株[5]。这种新型双病毒载体共同介导的编辑系统无须通过遗传转化即可获得后代编辑材料,进一步简化了病毒诱导基因编辑系统的操作。
除上述病毒诱导的基因编辑系统外,大麦条纹花叶病毒(barley stripe mosaic virus,BSMV)的gRNA递送系统可用于小麦和玉米中CRISPR/Cas9介导的靶向诱变,对小麦和玉米中的TaGASR7和ZmTMS5基因分别实现了78%和48%的基因编辑效率[64]。甜菜坏死黄脉病毒(beet necrotic yellow vein virus,BNYVV)gRNA 递送载体在转基因Cas9 本氏烟中对NbPDS3的编辑效率高达85%[65]。将CRISPR/Cas9 和sgRNA 插入到负链RNA 病毒苦苣菜黄网弹状病毒(sonchus yellow net rhabdovirus,SYNV),可实现90%~100%的编辑效率[6]。苹果潜隐球形病毒(apple latent spherical virus,ALSV)同样成功地对2个大豆GW2同源基因进行了编辑[66]。
虽然TRV 和PEBV 病毒能够侵染植物分生组织,但多数病毒仍无法进入植物分生组织,可遗传的基因突变效率很低。研究人员通过使用核糖体蛋白S5A(ribosomal protein S5A,RPS5A)启动子驱动SpCas9 表达并获得转基因株系,通过TRV 递送sgRNA 进入烟草叶片,后代种子有2%~6%发生了基因突变[67]。另外,通过在sgRNA上添加促进细胞间移动的信号,例如开花基因座T(Flowering Locus T,FT)、转运RNA(transfer RNA, tRNA)等,利用TRV 递送带有移动信号的sgRNA 至过表达Cas 转基因烟草中,后代有65%~100%的突变是可遗传的[68],该方法与通过传统的组织培养获得的突变体相比,大大缩短了时间、节省了经济和人力成本。
4 展望
CRISPR/Cas 基因编辑技术的出现彻底颠覆了整个生命科学研究领域,在短短几年间,CRISPR/Cas基因编辑技术已经成为领域内最受关注的研究工具。该技术也为植物性状改良、抗性提升,尤其为抗病毒育种工程研究开辟了新途径、提供了新思路。
目前,CRISPR/Cas基因编辑技术已被成功应用于多种抗病毒研究中,然而该技术还存在一些局限性,例如使用CRISPR/Cas9 工具抵抗外源病毒侵染时,双生病毒可以逃避CRISPR/Cas9 介导的抗性。因此,CRISPR/Cas9 能否加速双生病毒进化仍是一个需要探讨的问题。此外,通过病毒递送CRISPR元件受限于病毒包裹外源蛋白的容量,仅有SYNV和PVX 能够表达Cas 蛋白,因而开发较小分子量Cas 蛋白以用于病毒载体传递具有重要的应用价值。近日,有2个团队分别开发出一个新型的“迷你版”CRISPR/Cas12f1 的Un1Cas12f1 和AsCas12f1,大小仅为CRISPR/Cas9的一半,但编辑效率与Cas9相当[70]。利用小分子量蛋白进行病毒诱导的基因编辑系统的开发和应用可以完全避免转基因操作,消除外源蛋白对寄主的潜在影响,为未来作物育种研究提供更安全的种质资源。
此外,已有研究多基于提升寄主植物对病毒的抵抗力或直接靶向病毒序列来达到抗病毒的目的。但有一大部分植物病毒是将昆虫作为媒介进行传播,因此,能否通过同时控制昆虫介体的传播效率和增强植物对病毒的抵抗能力来控制病毒的侵染、复制、转录等多个过程,达到更为高效的抵抗病毒的效果,获得更广谱的抗病毒新材料仍是需要探索的方向。病毒诱导的基因编辑能够绕开最传统的组织培养方法来获得转基因材料,但多数病毒无法进入植物的分生组织,虽然有研究报道,在gRNA上添加移动信号能够实现遗传突变,但其后代编辑效率相对较低,因此亟须开发出更高效、稳定、便捷、可遗传的基因编辑系统,拓宽病毒介导的基因编辑系统的应用范围,更好地利用庞大的植物病毒王国为科学研究作出更多贡献。
猜你喜欢
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
中国瓜菜(2020年8期)2020-09-26
中成药(2018年7期)2018-08-04
现代装饰(2018年3期)2018-05-22
火花(2016年7期)2016-02-27
中国艺术时空(2014年2期)2014-02-28
