
雷公山保护区甜槠群落乔木层物种多样性及其相关性研究
2023-01-11 13:09:16叶荣欢李登江余德会郑德谋谢镇国吴必锋古定豪
湖南林业科技 2022年6期
叶荣欢, 李登江, 余德会, 郑德谋, 谢镇国, 吴必锋, 古定豪
(贵州雷公山国家级自然保护区管理局, 贵州 雷山 557100)
物种多样性海拔分布格局始终是生物多样性研究中的热点问题[1-2]。α和β多样性是物种多样性研究中的两个常用指标,其中α多样性是用于测量群落内生物种类数量以及生物种类间相对多度的一种测量,是表征植被群落内物种组成特征的一项重要指标,β多样性则度量群落的物种组成沿环境梯度的周转速率,即群落间相异性,被用于表征植被群落间物种组成差异,而海拔梯度被认为是影响α多样性与β多样性决定性因素之一[1-5]。
甜槠(Castanopsiseyrei)是属壳斗科(Fagaceae) 栲属(Castanopsis)常绿阔叶乔木,冠幅庞大,树形优美,林相整齐,广泛分布于我国长江流域以南的省份,常与拷属其他种及樟科树种伴生,是贵州省雷公山常绿阔叶林的典型代表[6-9]。目前,对于雷公山甜槠群落主要集中在生物量[7,10]、物种结构与多样性特征[11]方面的研究。关于雷公山甜槠群物种多样性在海拔梯度变化未见报道。本文以雷公山甜槠群落为研究对象,综合调查雷公山甜槠群落物种组成与物种多样性沿海拔梯度的变化,旨在深入了解雷公山甜槠群落物种多样性的海拔分布格局,为雷公山甜槠群落生物多样性保护和管理提供依据。
1 研究区概况
雷公山国家级自然保护区地处贵州省黔东南苗族侗族自治州境内,地理坐标为108°09′—108°22 E,26°15′—26°22′ N,总面积47 300 hm2。是以台湾杉(Taiwaniacryptomerioides)等珍稀动植物资源及其生态环境为主要保护对象,具有综合经营效益的亚热带山地森林生态系统类型的自然保护区[12]。雷公山保护区山地土壤垂直带明显,从海拔650~1 400 m为山地黄壤,1 400~2 000 m为山地黄棕壤,2 000 m以上分布有山地灌丛草甸土和山地沼泽土[13]。年降雨量1 300~1 600 mm,降雨量较多,属中亚热带山地季风湿润气候,植被类型为中亚热带常绿阔叶林,具有显著的垂直差异和坡向差异,独特的地理位置和优越的自然条件为多种多样的生物物种生长发育提供了良好的生态环境[7,14-15]。
2 研究方法
2.1 样地设置与调查方法
2021—2022年,在雷公山保护区采用样地调查法对海拔680~1 200 m,土壤类型均为黄壤的区间,选取具有代表性的甜槠群落设置一个面积为600 m2(20 m×30 m)的样地,每个样地设置10个样方(6 m×10 m)对甜槠群落乔木层进行调查。调查内容包括物种名、树高、胸径、冠幅、环境因子等(见表1)。
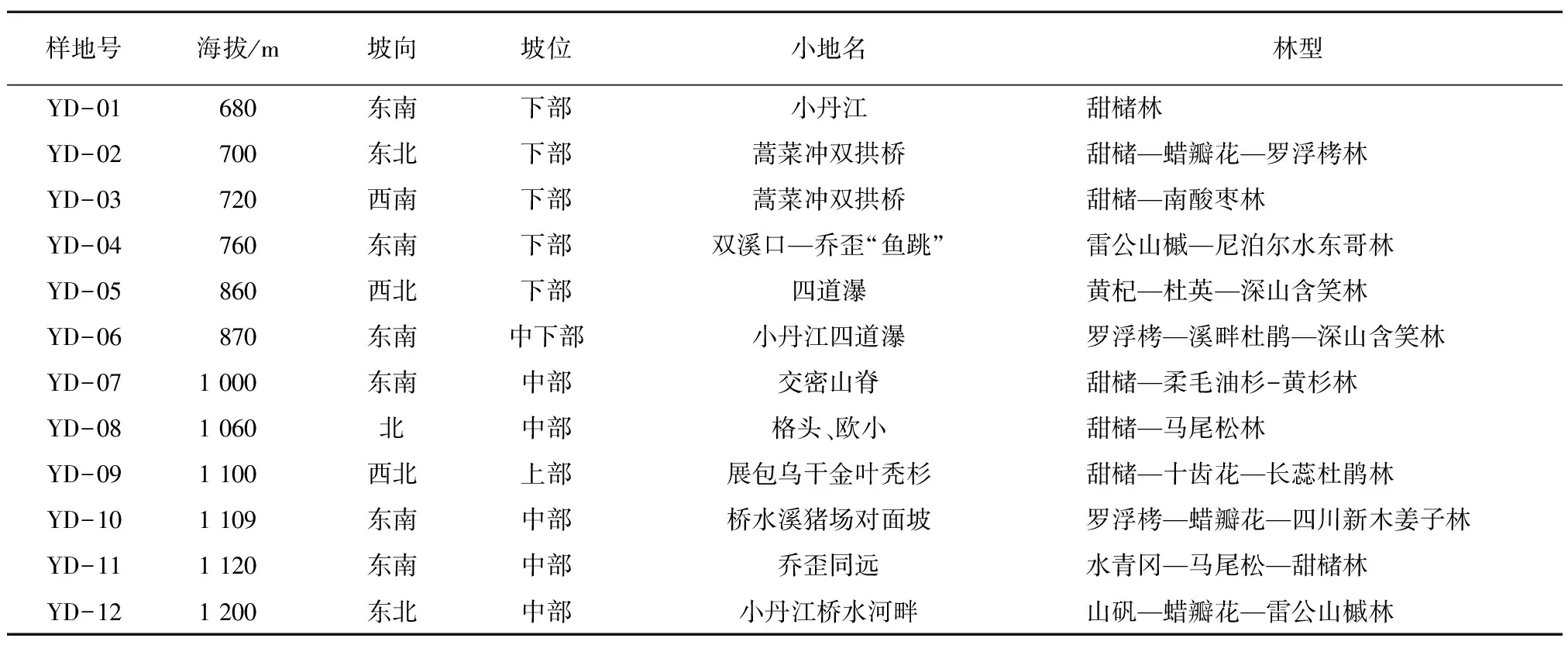
表1 样地概况
2.2 数据处理及计算方法
调查数据处理与统计分析采用 SPSS 19.0 软件和Excel 2010 软件完成。
2.2.1 重要值[16]
重要值=相对多度+相对频度+相对显著度
(1)
2.2.2 群落α多样性[1,16]
物种丰富度指数(D1)Gleason 指数:
(2)
物种丰富度指数(D2)Marglef 指数:
(3)
Shannon-Wiener 物种多样性指数(SW):
SW=-∑Pilnpi
(4)
修正后的 Simpson 生态优势度指数(SN):
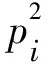
(5)
种间机遇机率 Hurlbert 指数(H):
(6)
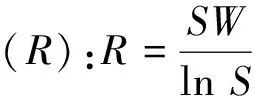
(7)
式中,S为研究系统中物种总数;A为单位面积;pi为种i的相对多度;N为所在群落的全部种的个体总数。
2.2.3 群落β多样性测定[1,16]
Whittaker指数(βw):
(8)
式中,S为研究系统中的物种总数;ma为各样方中物种的平均数。
Cody指数(βC):
(9)
式中,g(H)为沿海拔梯度上的新增种群数;l(H)为沿海拔梯度上失去的物种数目,即在上一个梯度中存在而在下一个梯度中没有的物种数目。
Routledge指数(βR):
(10)
式中,S为研究系统中的物种总数;c为两个样方(地)共有的物种数目。
2.2.4 群落相似性
采用 Jaccard 相似性公式计算相似性系数[1,16]:
(11)
式中,q为群落相似性系数;c代表两个样方共有的物种数目;a和b分别代表两个群落的物种总数。
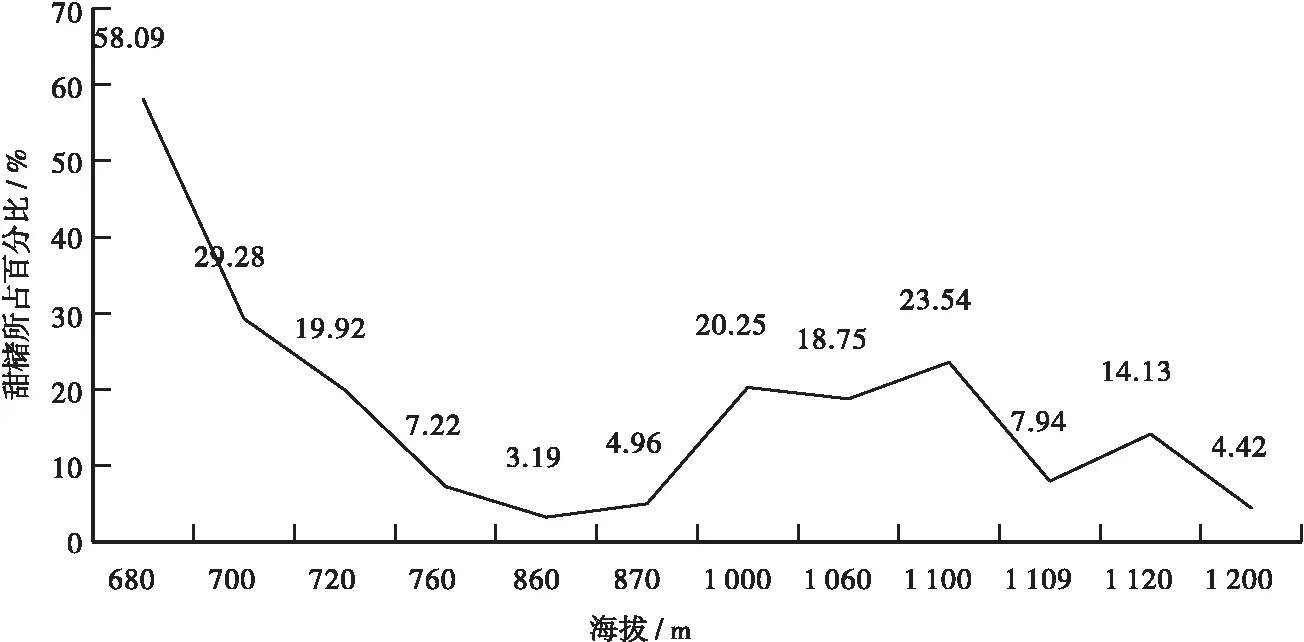
0.00 甜槠群落乔木层中共有维管束植物40科79属143种,其中,裸子植物有3科6属8种;双子叶植物有37科73属135种。由此可知,在雷公山保护区分布的甜槠群落乔木层中双子叶植物的物种数量占据绝对优势(见图1)。 图1 甜槠群落乔木层物种组成Fig.1 Species composition of tree layer of Castanopsis eyrei community 从甜槠重要值在群落乔木层中所占百分比可知,甜槠的重要值在海拔680 m、700 m、720 m、1 000m、1 060 m、1 100 m样地中,分别为58.09%、29.28%、19.92%、20.25%、18.75%、23.54%,位居第一,为优势种群,在760 m、860 m、870m、1 109 m、1 120 m、1 200 m样地中分别排在第4(7.22%)、第8(3.19%)、第6(4.96%)、第4(7.94%)、第3(14.13%)、第10(4.42%),没有占据优势;沿海拔梯度来看,甜槠的重要值表现出随着海拔的升高而降低的总体趋势(见图2)。 3.2.1 α多样性沿海拔梯度的变化 甜槠群落乔木层 α 多样性结果表明,物种多样性指数与生态优势度指数、种间机遇指数与群落均匀度、丰富度 Gleason 指数与丰富度 Marglef 指数沿海拔梯度的变化分别表现出一致性,物种多样性指数与生态优势度指数在680~870 m表现为先升高后降低,在870~1 200 m表现为先升高后平缓、再降低又升高的趋势;种间机遇指数与群落均匀度的变化趋于平缓;丰富度 Gleason 指数与丰富度 Marglef 指数在680~860 m先剧烈升高剧烈降低后、又升高再降低,在860~870 m中趋于平缓,在870~1 200 m表现为先升高再降低、并趋于平缓后又升高的趋势(见表2)。 图2 甜槠所占百分比随海拔的变化Fig.2 Change of percentage of Castanopsis eyrei with altitude 表2 甜槠群落 α多样性指数 这一结果与随着海拔高度的增加,群落物种多样性逐渐降低的大多数情况不同,可能是选取的样地海拔梯度较小,海拔没有成为限制植物分布的主要因素,样地也受到一定人为干扰的原因[2]。 3.2.2 α 多样性指数间的相关性及回归分析 根据Pearson 相关性分析结果可知,物种多样性指数、生态优势度指数与其他指数存在P<0.01水平上显著正相关,种间机遇指数与群落均匀度在P<0.01水平上显著正相关,丰富度 Gleason 指数与丰富度Marglef 指数在P<0.01水平上显著正相关(见表3)。 表3 甜槠群落 α 多样性指数间相关系数Tab.3 The correlation coefficient between α diversity index of Castanopsis eyrei community. 由于物种多样性指数跟其他指数之间存在P<0.01水平上显著正相关,对物种多样性指数与其他各指数之间进行了回归分析: 物种多样性指数与生态优势度指数的回归方程: Y=0.717+0.838X (12) 物种多样性指数与种间机遇指数的回归方程: Y=-0.935+3.931X (13) 物种多样性指数与均匀度指数的回归方程: Y=-0.428+3.662X (14) 物种多样性指数与丰富度 Gleason 指数的回归方程: Y=1.265+0.354X (15) 物种多样性指数与丰富度 Marglef 指数的回归方程: Y=1.321+0.354X (16) 从对物种多样性的率贡献上看,生态优势度对群落的多样性指数贡献率最高,从高到低为生态优势度指数>种间机遇指数>丰富度指数>群落均匀度指数。 3.2.3β多样性沿海拔梯度的变化 从表4可知,甜槠群落乔木层β多样性沿海拔梯度的变化基本呈波状,沿海拔梯度Whittaker指数的变化不明显,Cody指数与Routledge指数的变化基本一致,并没有表现出一定的规律性;其中Cody指数与Routledge指数分别在760 m、1 100 m处出现极值,说明甜褚群落乔木层在这两个海拔区间的物种组成差异较大。这一结果表明β多样性指数在海拔梯度较小时,受相邻样地间地形因子与水平距离的影响较大,其波动大小与相邻样地间的地形因子、水平距离引起的物种构成变化大小有关[2,18]。 表4 甜槠群落 β 多样性指数Tab. 4 Species β diversity index of Castanopsis eyrei community 群落相似系数是对各群落间物种共有程度的度量。如表5所示,海拔760 m与海拔1 109 m群落相似性系数为0.25,呈中等不相似水平,其他样地之间的相似性系数均在0.00~0.25之间,处于极不相似水平,说明各样地间物种组成存在较大的差异,物种多样性明显;沿海拔梯度来看,随着海拔差距的增大,群落相似性系数呈逐渐减小的趋势,说明海拔差异对相似性系数呈负效应,反映了与海拔相关的环境因子于群落构建的重要作用[2];从不同海拔样地来看,相似性系数最大的0.25,最小的0.03并没有出现在海拔高度相近与海拔高度差距较大的样地之间,说明在海拔变化不明显时,地形因子与水平距离对生境及群落结构差异性的影响较大。 表5 不同海拔甜槠群落相似性系数 (1) 甜槠群落乔木层在雷公山海拔680 m~1200 m的物种组成差异大,优势种随着生境的变化而发生改变,说明甜褚群落乔木层物种组成受海拔、地形等多因素控制。 (2) 雷公山保护区甜槠群落乔木层α多样性指数沿海拔梯度的变化除种间机遇指数与群落均匀度指数平缓外,其他指数的变化不规律。表明在地海拔梯度较小时,海拔没有成为限制植物分布的主要因素,群落的物种组成与生境受地形变化的影响较大[2]。 (3) 甜槠群落乔木层物种多样性指数与其他指数存在P<0.01水平上显著正相关。在整个甜槠群落乔木层中生态优势度对群落的多样性的贡献率最高,从高到低为生态优势度>种间机遇>丰富度>群落均匀度。 (4) 甜槠群落乔木层β多样性沿海拔梯度的变化大体呈波状,除Whittaker指数外,其他指数沿海拔梯度在760 m和1 100 m处出现极值。说明在雷公山保护区海拔680~1 200 m内,垂直空间距离对β多样性的作用没有水平空间距离与地形因子的作用大;垂直空间尺度较小时,扩散限制等过程对于甜槠群落乔木层物种组成的影响不明显。 (5) 不同海拔甜槠群落乔木层的相似系数除在海拔760 m与海拔1 109 m群落相似性系数为0.25,呈中等不相似水平,其他样地之间的相似性系数均在0.00~0.25之间,处于极不相似水平。说明雷公山保护区拔甜槠群落乔木层在海拔600~1 200 m之间物种多样性明显,相邻海拔的生境及群落结构差异性受地形因子与水平距离影响较大,相似系数沿海拔的变化反映了与海拔相关的环境因子于群落构建的重要作用。3 结果与分析
3.1 甜槠群落结构特征

3.2 甜槠群落沿海拔梯度的物种多样性



3.3 不同海拔甜槠群落相似性系数

4 结论与讨论
猜你喜欢
数学物理学报(2022年5期)2022-10-09 08:56:44
建筑与预算(2022年6期)2022-07-08 13:59:18
河北画报(2020年8期)2020-10-27 02:54:20
农民致富之友(2018年23期)2018-12-19 14:58:54
吉林农业(2016年12期)2017-01-06 19:41:53
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:51
原生态民族文化学刊(2014年1期)2014-08-08 14:46:40
中央民族大学学报(自然科学版)(2014年2期)2014-06-09 08:28:24
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
