
我国城市鸟类多样性特征及其影响因素研究*
2022-12-20 02:32:12田永莲孙秀锋吕鲜艳李旭
西南大学学报(自然科学版) 2022年12期
田永莲, 孙秀锋, 吕鲜艳, 李旭
西南大学 园艺园林学院, 重庆 400715
城市生物多样性保护对于维护城市生态安全具有重要意义[1], 城市鸟类是城市生物多样性保护的主要对象和城市生态系统的重要组成部分[2-3], 在生态系统中鸟类是生态金字塔顶端的上层物种, 对栖息地的组成和环境变化非常敏感, 因而也是城市生态环境和城市生物多样性的重要指示物种[4], 可以发挥城市状况晴雨表的作用[5]. 此外, 由于鸟类在城市环境中分布广泛且易于接近, 是居民在城市环境中亲近自然的重要媒介, 具有娱乐和教育等社会功能[6], 对城市居民的身心健康和人类福祉有重要作用[7], 因此城市鸟类对城市生态系统健康具有重要意义.
正是由于对栖息地变化的异常敏感性, 使鸟类成为城市化过程中最受干扰的物种之一[4, 8]. 在城市化高速发展压缩其生存自然生境的同时[9], 城市景观岛屿化、 破碎化、 高异质性及多边界等特征, 导致城市鸟类在组成、 结构、 分布及营巢方式等方面发生显著变化[10-11], 如鸟类多样性降低[12]、 食性类型结构改变[13]、 本地鸟类减少而外来鸟类种类增加[14]、 不同城市鸟类同质化等[15]. 目前, 针对我国城市鸟类多样性特征, 学者们多关注于对单个城市鸟类组成的研究[16-17], 或生境类型对城市鸟类种数的影响等[18-19], 表现为重视中观层面的城市尺度而忽视宏观层面上生物多样性的研究[1], 尤其是鲜见对不同城市之间城市鸟类组成和多样性比较及整体性特征的研究. 我国是全球12个生物“巨大多样性”(Mega-diversity countries)国家之一[20], 拥有1 371种鸟类, 约占全球鸟类种数的14%[21]. 经过后续鸟类的自然增补, 目前我国拥有1 471种鸟类. 尽管目前在一些城市鸟类物种多样性与城市生境关系研究方面取得了较大的进展, 并在城市环境因子对鸟类的影响方面发现了一些一般性规律[22-23], 但进一步从总体上厘清我国城市鸟类多样性分布与构成特征并分析其影响因素, 对于城市生物多样性保护及城市环境可持续发展具有重要意义.
本研究以21个城市鸟类数据为基础, 分析我国城市鸟类多样性特征, 从居留类型、 食性类型等方面分析其组成特点, 并进一步将城市自然条件、 社会经济等因子作为环境解释变量, 解析城市鸟类多样性和组成特征与相关驱动因子之间的关系, 探寻我国城市鸟类多样性和组成分布格局, 以期为城市化过程中生物多样性保护提供有益参考.
1 数据与方法
1.1 数据来源
本研究中城市鸟类数据主要来源于文献、 相关数据库和实地调研, 通过关键词“城市鸟类” “城市鸟” “市区鸟类” “城市鸟多样性” “Urban birds” “Urban bird diversity”等分别在中、 英文学术数据库中检索与中国城市鸟类相关的论文, 在符合条件的80篇文献中包含有“鸟类名录”的共计31个城市, 其中某些城市仅包含了单一季节的鸟类, 本研究对该类城市进行剔除, 确定了21个城市作为鸟类统计对象, 其中哈尔滨市鸟类调查数据来源于实际调查结果[24]. 为了使研究数据更加完善, 本研究通过“鸟人课堂”[25]和“中国观鸟记录中心”[26]对21个城市鸟类数据进行了补充, 数据获取时间为2019年12月. 为了避免不同文献或记录人员对同种鸟使用不同名称导致重复现象, 依据中国观鸟年报《中国鸟类名录6.0》[27]、 《中国鸟类分类与分布名录》[21]及《中国生物物种名录2020版》[28]对鸟类名称进行统一, 并确定了其目、 科、 属.
为了分析城市鸟类组成的影响因素, 本研究选取了城市区位及8个自然和社会因子作为分析指标. 城市自然环境因子数据主要选取海拔、 年均温度、 年均降雨量、 城市绿化覆盖度、 绿度植被指数(GVI)及植物丰富度等因子, 除海拔外其余数据均来自世界气候网站[29]; 城市社会经济因子数据主要选取人口密度、 城区面积作为分析因子, 数据来源于《中国城市统计年鉴》[30].
1.2 数据收集及分析方法
本研究选取的21个城市具有不同的规模等级, 并分布于我国7个鸟类亚区, 在一定程度上可以代表不同类型城市和分布区域特征. 根据上述方法统计获得4 234个城市鸟类数据, 分布于27目92科. 影响因子可分为3类, 区位、 城市自然因子及社会经济因子, 其中城市空间距离因子为各城市经、 纬度数据转换后的空间距离(表1).
以收集的21个城市鸟类科(属、 种)、 居留类型以及食性数据为基础, 采用数据透视表的形式分析城市鸟类的组成特征, 并将城市鸟类科(属)内物种数量频次与全国鸟类科(属)内物种数量频次进行比较及聚类分析, 以揭示城市鸟类多样性特征. 然后, 用单因子回归方法进行城市鸟类多样性影响因素分析. 为进一步厘清城市鸟类多样性组成差异的影响因素, 本研究将鸟类多样性进行分解(POD法)[31], 并分别对其进行了Mantel检验分析.

表1 城市自然及社会经济因子
2 结果与讨论
2.1 我国城市鸟类多样性分析
2.1.1 城市鸟类多样性概况
21个样本城市共记录城市鸟类763种, 占中国鸟类种数的51.87%, 隶属于27目, 92科, 289属(表2). 本研究发现各鸟类种数在各科间分布极不均衡, 92科中有28科仅有1个种类, 而种类最多的10个科中分布有332个种, 占全部种数的43.51%, 种数排名前3的是鹟科(Muscicapidae)、 鸭科(Anatidae)以及鹬科(Scolopacidae), 其中鹟科有68个种数占城市鸟类统计总种数的8.91%, 鸭科有36种数占4.72%, 鹬科有35个种数占4.59%. 从城市鸟类构成优势度来看, 鸭科鸟类优势度最强, 21个城市中以鸭科为优势科的城市有10个, 占城市数量的47.62%, 但均为北方城市; 鹟科鸟类紧随其后, 作为优势科的城市占33.33%; 而以鹬科为优势科的城市仅占9.52%. 由此可见, 鸭科和鹟科鸟类在城市鸟类群落种类组成中占主要地位; 同时有195种鸟类仅在一个城市出现, 即21个城市中平均每个城市约有9种鸟类是独有鸟类, 各城市间鸟类多样性组成差异较大. 从种的分布广度来看, 白鹡鸰(Motacillaalba, 鹡鸰科)、 家燕(Hirundorustica, 燕科)及麻雀(Passermontanus, 雀科)3种鸟类分布最为广泛, 在21个城市中均有分布; 其次是黑水鸡(Gallinulachloropus, 秧鸡科)、 远东山雀(Parusmajor, 山雀科), 属于分布第二广泛的鸟类.
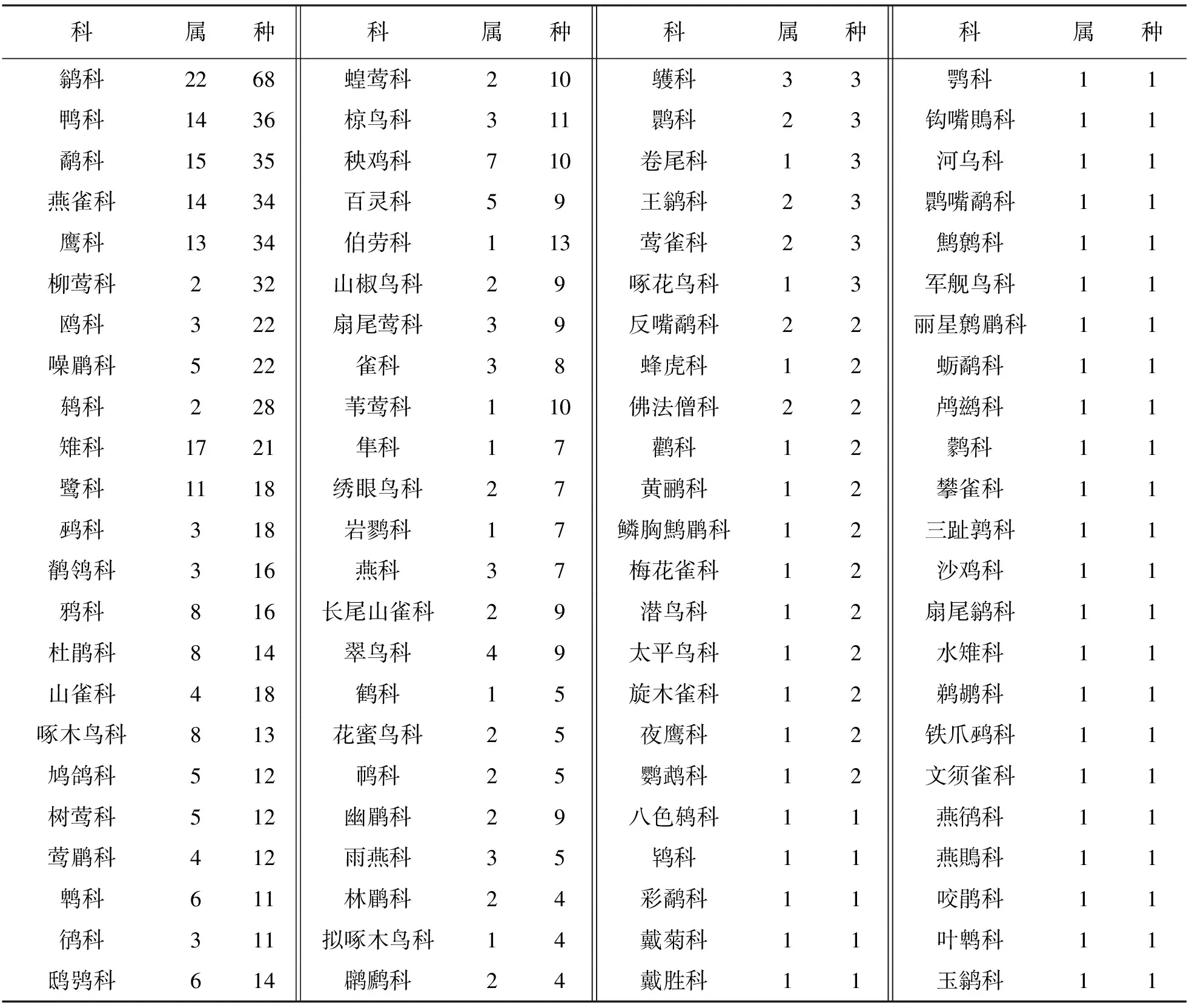
表2 21个城市鸟类各科、 属、 种数量
2.1.2 城市鸟类多样性特征
已往的研究表明, 物种与高级分类阶元在数量上密切相关[32], 物种与科(属)的数量关系和频次分布规律在一定程度上反映了物种多样性的特点. 为解析城市鸟类多样性特征, 本研究将城市鸟类种数和全国鸟类种数分别与科(属)频次分布规律进行对比(图1).
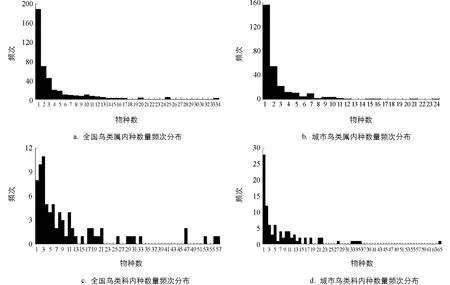
a,c引自参考文献[32], b,d为作者自绘.图1 全国鸟类与城市鸟类属(科)内物种数量频次分布对比
通过比较发现, 城市鸟类属间种数频次呈非均匀分布, 随着物种数量增多频次降低, 呈幂模型状态分布, 与全国鸟类属间物种数量频次分布规律基本一致. 城市鸟类科间种的数量频次分布与全国鸟类科间种的数量频次分布也呈相似状态, 随物种数量增多频次逐步降低, 但波动性比属间频次分布波动性大. 不同的是, 虽然两者都是鸟类种数小于3的科频次最高, 但是从全国来看频次最高的是有2~3个种类的科, 而城市鸟类频次最高的是只有1个种类的科, 且显著高于其他频次, 说明在城市鸟类中有更多的科仅有1个科内种在城市区域被发现, 即城市中鸟类物种的丰富性更加脆弱, 如果这个鸟类物种迁出或消亡, 则代表相应的科类也会在城市地域消失, 对城市鸟类多样性的影响更大.
2.1.3 不同城市鸟类多样性特征差异
从城市鸟类种类多样性差异来看, 不同城市鸟类种类数量差异明显, 以上海为代表的排名前5的城市平均鸟类种数达320种, 而排名后5的城市平均仅有97种, 分析发现城市区位与城市规模在其中发挥了一定的作用, 符合生产力假说[33]及面积效应. 为进一步分析不同城市鸟类多样性特征差异, 本研究将21个城市鸟类物种与高级分类阶元数量关系进行聚类分析(图2), 发现呈明显的4个类型. 进一步统计分析这4个类型的属科比、 种属比及种科比, 发现分别呈现出低-低-低、 高-低-低、 高-低-高及高-高-高的特征. 临沂等10个城市为低-低-低型, 该类城市不但鸟类多样性较低, 平均仅有115种, 而且对应每科或属中的种类多样性也低, 说明每种城市鸟类对维护科或属水平上的多样性压力更大; 北京等4个城市为高-高-高型, 表明该类型城市不但鸟类物种丰富度高, 平均有315种, 而且对应每个科内属及属内种也多, 整体反映了该类型城市鸟类多样性较高; 而杭州等4个城市为高-低-低型; 广州等3个城市为高-低-高型; 它们的鸟类多样性也呈现出各自的特点. 但总体来看, 低-低-低型城市最多, 约占研究城市数量的50.00%, 表明我国城市鸟类多样性保护整体形势严峻, 也反映出不同城市之间地域差异带来了极大的城市鸟类多样性差异.
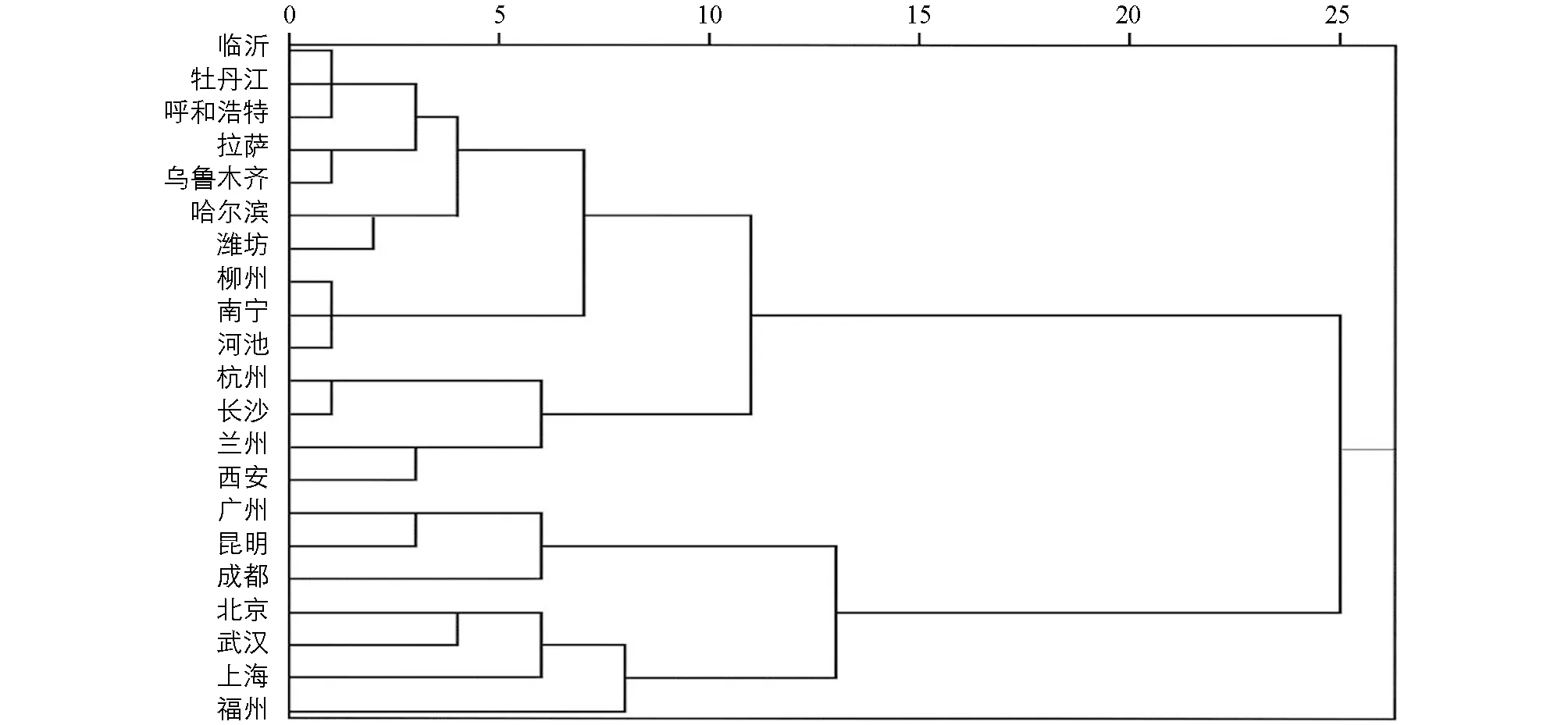
图2 城市鸟类物种与高级分类阶元数量关系型聚类
2.2 城市鸟类居留及食性类型构成特征
2.2.1 居留类型构成特征
鸟类活动范围巨大且不同类型鸟类差异明显, 根据居留特征可分为留鸟及候鸟等类型. 我国领土跨越古北界和东洋界鸟类区系, 是世界上包含两个动物地理界的仅有两个国家之一[34], 且处于世界候鸟南北、 东西迁徙通道较为关键的位置, 全球8条候鸟迁徙通道中有3条与我国鸟类迁徙密切相关[35]. 因此, 了解我国城市鸟类居留类型构成特征, 对鸟类多样性保护具有重要意义. 本研究分析得知, 我国城市鸟类居留类型由多到少依次为留鸟、 夏候鸟、 冬候鸟及旅鸟(表3). 其中, 留鸟有424种, 占城市鸟类种数的55.57%, 其次夏候鸟315种, 占41.28%. 每种类型的空间分布符合我国城市在生物地理区中的区位, 如在21个城市中47.61%的城市是以留鸟为主要城市鸟类, 除了西安和兰州外, 其余城市均为南方城市, 其中昆明、 南宁及广州等城市尤为明显; 其次有28.57%的城市以夏候鸟为主要城市鸟类, 这些城市均为北方城市, 其中哈尔滨尤为显著; 上海、 武汉等亚热带地区城市则以冬候鸟为主要类型. 值得注意的是, 北京和成都的旅鸟类型所占比例最大, 这与城市处于鸟类重要迁徙通道有很大关系. 因此, 城市鸟类多样性保护应充分考虑城市所在的生物地理区, 根据居留比例及居留特征制定相应的鸟类多样性保护策略.
2.2.2 食性类型构成特征
从食性角度对城市鸟类构成进行分析, 有助于了解城市鸟类内部资源利用及种间共存关系, 并能从侧面反映城市自然环境状况. 根据食性特征可将城市鸟类划分为杂食性、 植食性、 食鱼性、 食肉性及食虫性几种类型. 总体来看, 21个城市鸟类中食性种类数量由多到少依次为食虫性、 杂食性、 食鱼性、 食肉性及植食性鸟类(表3). 食虫性鸟类种数最多, 占统计总种数的39.84%; 其次杂食性鸟类占统计总种数的37.75%; 而最少的植食性鸟类, 仅占统计总种数的4.06%. 这是因为食虫性鸟类相较而言获得食物的能力更强, 杂食性鸟类取食种类更丰富, 可以在城市中良好地适应[36], 而城市中自然植被减少, 导致植食性鸟类最少. 从城市差异来看, 61.90%的城市以食虫性鸟类为主要类型, 主要为分布在横断山脉以东、 秦岭淮河以南地区的城市, 比例最高的是河池, 与其相近的柳州、 南宁及云南省的昆明也位居前列, 这是因为这些城市处于亚热带地区, 城市植被丰富性高, 为食虫性鸟类提供了良好的食物资源及栖息地; 北京、 哈尔滨、 拉萨等7个城市以杂食性鸟类为主, 占统计城市的33.33%, 主要分布在华北、 东北、 西部等地区, 比例最高的是乌鲁木齐, 这可能是因为典型的西北内陆自然环境中单一取食的鸟类本来较少, 而城市环境中的城市适应种相对聚集较多形成的, 其中西安与呼和浩特两个城市的杂食性鸟类和食虫性鸟类比例相当; 食鱼性鸟类比例最高的是上海, 这与城市及周边水网密布湿地资源丰富有关; 食肉性鸟类比例最高的是拉萨, 这与拉萨鸟类总数相对较少, 而鹰科、 隼科等鸟类相对较多有关.
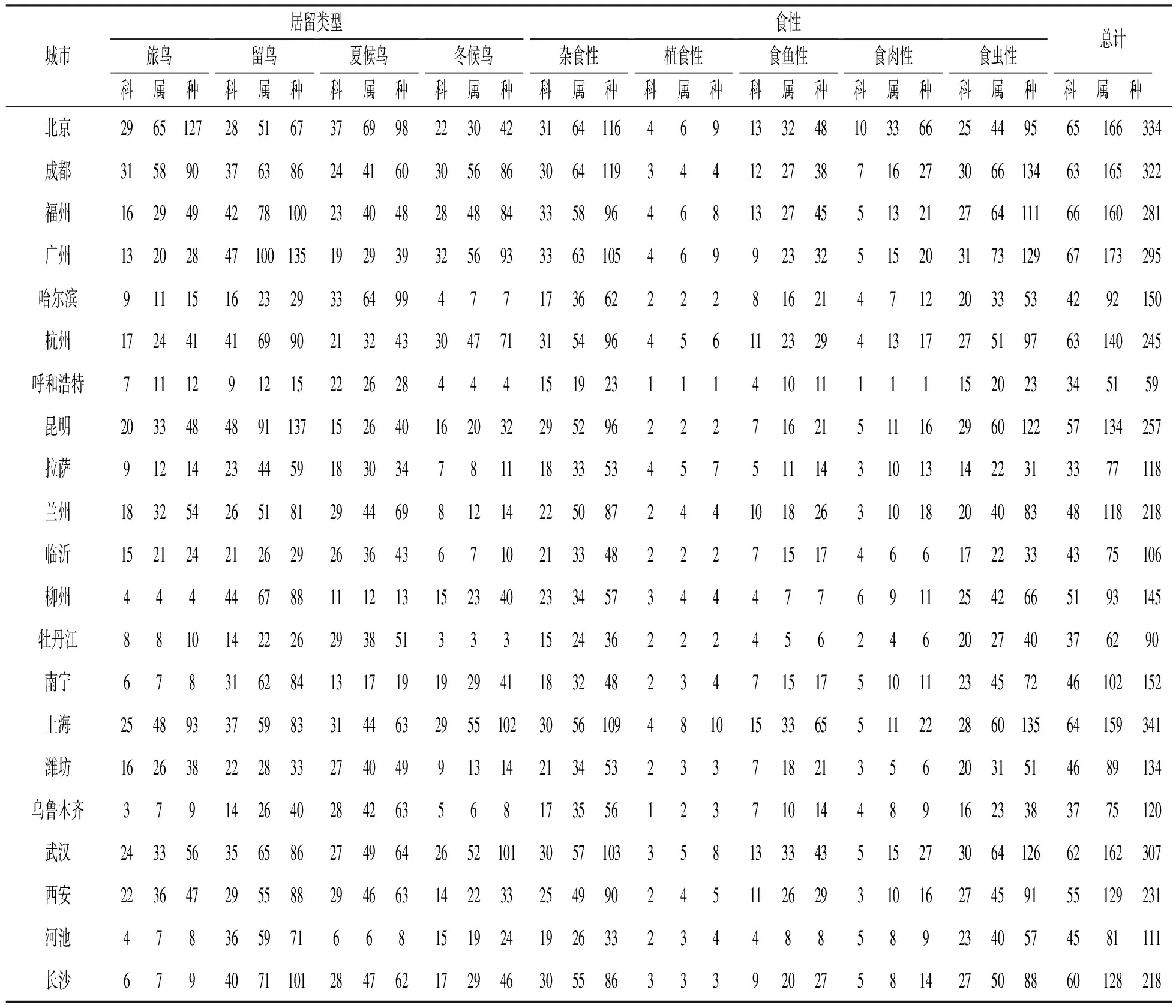
表3 中国21个城市鸟类组成各科、 属、 种表
2.3 城市鸟类多样性影响因素分析
2.3.1 城市鸟类多样性单因子分析
尽管目前的研究认为, 影响我国大尺度鸟类丰富度的环境因子主要为年降水量、 年均温和增强型植被指数(EVI)[37], 但城市鸟类有其构成的独特性, 如本地种灭绝而外来种增加[38], 鸟类构成同质化[13]等特点, 因此本研究选取经纬度、 高程、 绿度植被指数(GVI)等10个因子与城市鸟类多样性进行相关性分析, 发现从空间分布来看, 鸟类超过200种数的城市主要集中在东经102°~122°之间; 从纬度来看, 集中分布在北纬22°~35°及附近. 同时, 结果显示城市年均温度、 植物丰富度、GVI及城区面积与城市鸟类丰富度呈显著正相关关系(图3), 而对全国鸟类丰富度有显著影响的年均降水量因子[37], 对城市鸟类丰富度影响并不显著, 这应该是城市鸟类种类组成有别于自然区域, 造成城市鸟类对降水因素依赖度降低所致.
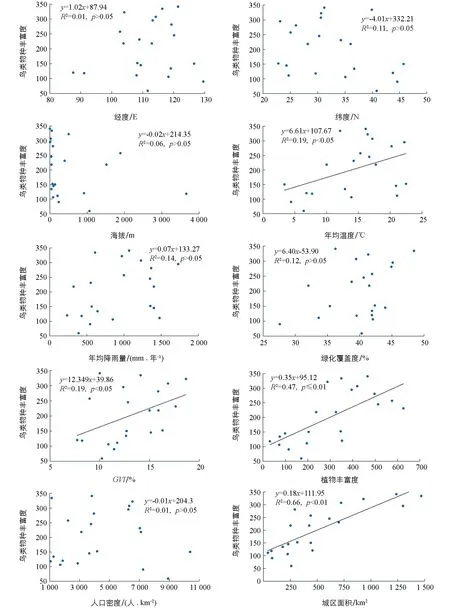
图3 城市鸟类多样性与相关因子一般线性回归散点图
2.3.2 城市鸟类多样性Mantel检验分析
为进一步分析城市鸟类多样性组成的影响因素, 应用Mantel检验进行影响因子分析. Mantel检验通过研究对象关联程度量化, 产生样本间两两距离矩阵, 并通过计算两个矩阵的相关性, 验证多元变量是否具有空间依赖性[39]. 为了揭示不同城市鸟类多样性组成差异的影响因素, 本研究同时应用POD法[40]将鸟类多样性进行分解. 不同城市鸟类种类组成的差异由物种替换和丰富度差异这两个过程决定[31], 在本研究中物种替换则表示一个城市的鸟类物种被另一个城市鸟类物种替换, 鸟类丰富度差异则表示各城市独有鸟类种数差值的绝对值. 将鸟类多样性组成、 替换及鸟类丰富度差异分别进行Mantel检验分析, 结果如表4所示.
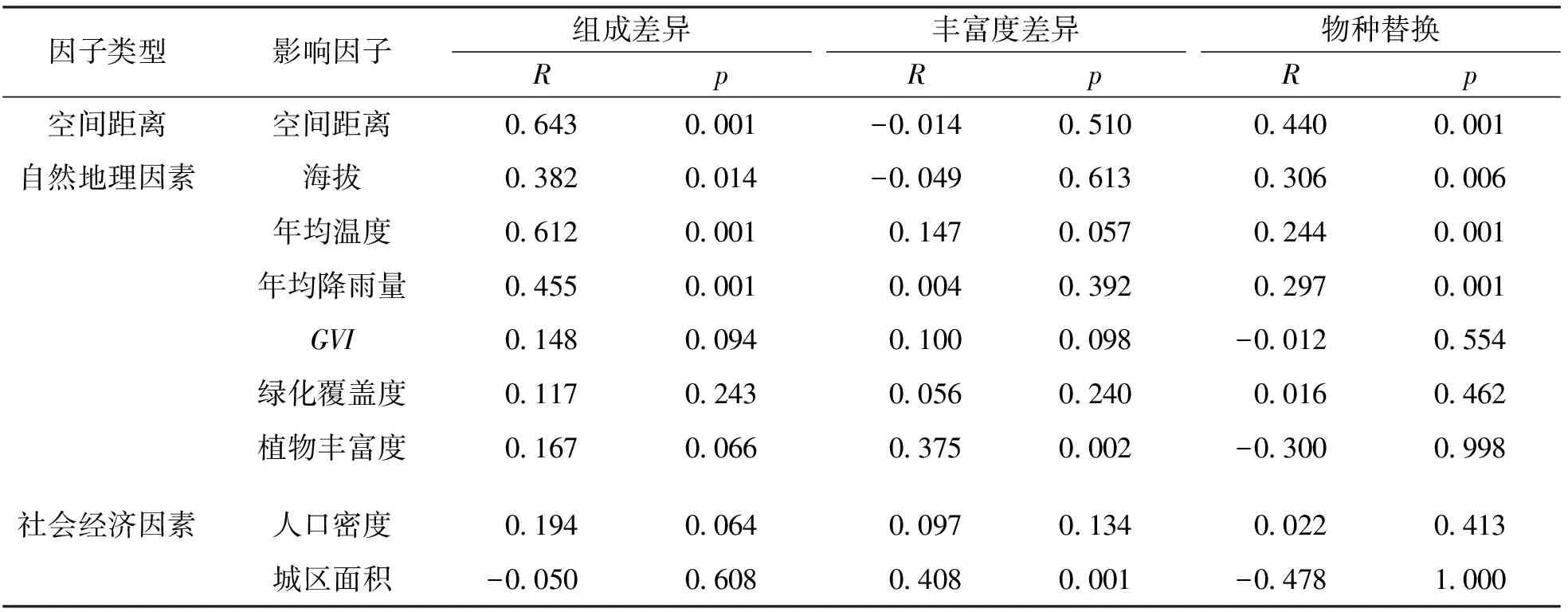
表4 城市鸟类多样性及其分解与各影响因子的Mantel检验分析
分析结果表明, 9个变量相加可以解释城市鸟类物种丰富度变异的约52.00%(调整后R2=0.021), 其中自然环境因素、 空间距离和社会经济因素互相独立(不相关、 正交), 即3者不存在共同解释的部分. 从城市鸟类多样性组成差异来看, 有显著关系的影响因子包括空间距离(R=0.643,p=0.001)、 年均温度(R=0.612,p=0.001)、 年均降雨量(R=0.455,p=0.001)及海拔(R=0.382,p=0.019)4个因子, 空间距离及海拔因子差异程度与城市自然环境条件相似性密切相关, 而温度及降雨量也属于自然环境因子, 因此可以看出造成城市鸟类组成差异的影响因子还是以水热等自然因子为主导, 符合生物多样性环境热量假说[33]和生产力假说[41]; 其中丰富度差异方面, 城区面积(R=0.408,p=0.001)及植物丰富度(R=0.375,p=0.001)与城市鸟类多样性呈显著正相关, 面积越大则物种丰富度越高, 植物丰富度反应出的生境类型及食物越丰富, 特别是对食虫性及植食性鸟类多寡有明显的影响, 可见植物丰富度越高鸟类多样性亦越高; 物种替换反映的是不同城市鸟类种类的独特性, 即假设各城市鸟类的多样性程度一致, 那么与各城市间鸟类种类差别程度有显著关系的因子包括空间距离(R=0.440,p=0.001)、 海拔(R=0.306,p=0.006)、 年均温度(R=0.244,p=0.004)及年均降雨量(R=0.297,p=0.001), 与影响鸟类组成差异的影响因子一致, 但从影响程度来看, 年均温度的影响程度显著降低, 而海拔的影响则相对升高, 即城市相对独特的地理条件造成了城市鸟类物种的替换.
3 结语
生物多样性是维系生态系统稳定健康及发挥服务功能的基础, 人类活动是造成生物多样性下降的主要原因[42], 而城市恰恰是高强度人类活动与丰富生物多样性并存的区域[43], 因此城市鸟类是城市生物多样性保护的主要对象[2]. 本研究通过对21个城市的鸟类分析发现, 763种城市鸟类分布于92科289属, 占我国鸟类总种数的51.87%, 各科间种数分布极不均衡, 城市鸟类物种在高级分类阶元分布差异显著, 其科内种数较非城市地域更加单一, 聚类分析发现约50.00%的城市鸟类种科(属)比维持在低水平, 反映出城市鸟类多样性系统的脆弱性更加明显, 表明总体上我国城市鸟类多样性保护压力较大. 在居留类型构成方面, 留鸟与夏候鸟较多, 分别占城市鸟类总种数的55.57%和41.28%. 以留鸟为主要城市鸟类的城市占47.62%, 除西安和兰州外均为南方城市, 可能是该两个城市处在中纬度偏南地区且规模较大, 从而使杂食性的城市适应性留鸟较多. 而北京和成都的旅鸟种数所占比例最大, 与其处于我国两条重要鸟类迁徙通道有关. 在食性类型构成方面, 61.90%的城市以食虫鸟类为主, 主要分布在我国东南部区域. 33.33%的城市以杂食性鸟类为主, 主要分布于华北、 东北、 西部等地区. 城市食性构成有助于了解城市鸟类内部资源利用及种间共存关系, 并能从侧面反映城市自然环境状况.
影响城市鸟类多样性的因子不同于自然地域大尺度鸟类多样性的影响因子, 对我国鸟类整体多样性有显著影响的年降水量因子对城市鸟类多样性的影响却不明显, 这应该是因为城市鸟类种类构成有别于自然区域, 使其对降水因素依赖程度降低所致. 通过Mantel检验分析, 本研究发现城市自然环境差异是造成鸟类多样性差异的主要原因, 其中丰富度差异与面积效应及植物丰富度显著相关, 因此城市环境营建中的植物丰富度是鸟类多样性的重要支撑; 导致不同城市间鸟类替换的因素与鸟类多样性差别因素基本一致, 但是从影响程度来看, 年均温度的影响程度显著降低, 海拔因子的影响则相对升高, 反映出我国在空间上东西向3大阶梯的影响增大, 南北向纬度导致温度变化的影响降低.
城市鸟类多样性需要在多尺度上分析和认识, 鉴于目前的研究多聚焦于城市尺度或绿地尺度, 本研究在国土尺度上通过对不同城市间鸟类多样性的比较与分析, 从整体上找出了城市鸟类多样性的一些基本特征、 城市差异及影响因素, 有助于城市鸟类多样性保护, 提高城市生态服务供给水平. 由于我国城市数量多、 分布广、 类型多样, 本研究仅以数据可得的21个城市为样本, 分析结果是否能全面反映我国城市鸟类多样性的基本面貌, 还有待进一步深入探寻. 同时, 本研究仅对鸟类种类多样性进行了分析, 实际上鸟类生态价值及服务功能还体现在单一种的丰富性上, 同时不同鸟类珍稀程度与保护等级也有差异, 这些问题也需要进一步在城市间比较与分析.
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:18
学与玩(2022年9期)2022-10-31 02:54:08
文苑(2020年12期)2020-04-13 00:54:14
小太阳画报(2019年1期)2019-06-11 10:29:48
科学大众(中学)(2019年3期)2019-05-17 10:04:30
数理化解题研究(2018年34期)2018-12-27 08:30:04
汽车观察(2018年10期)2018-11-06 07:05:26
数理化解题研究(2018年16期)2018-07-12 02:46:56
小学生必读(低年级版)(2017年5期)2017-08-12 03:47:07
少儿科学周刊·少年版(2015年1期)2015-07-07 17:15:12
