
流溪河底栖硅藻群落组成及其对水环境的季节响应研究
2022-12-17 12:36卓全龙
广东水利水电 2022年12期
陈 慈,谭 琳,卓全龙
(1.广东省水文局广州水文分局,广州 510150;2.暨南大学生态学系/水生生物研究中心,广州 510632;3.广州市水务科学研究所,广州 510220)
1 概述
在南亚带河流中,水体水动力季节强,增强了理化指标的瞬时性和指示片面性[1]。底栖硅藻是单细胞的真核生物,参与碳、氮、磷元素的生物地球化学循环,是河流生态系统中的主要组成生物[2]。底栖硅藻营固着生活,繁殖周期短,对环境变化敏感而反应迅速,其现存生物群落能够较客观和全面地体现河流当前一段时期内的环境与生态状况。底栖硅藻种类繁多,不同种类对环境压力的耐受性存在差异[3],它们在空间上分布紧凑,在一小块底质上就能采集到有代表性的群落,能有效指示环境和生态的变化,底栖硅藻已被广泛用于河流生态健康状况评估。
流溪河流域大部分在广州市辖区内,与广州市民生活联系紧密,又称为广州人的母亲河,但其生态环境受人类活动的影响严重。流溪河流域属粤北山区与珠江三角洲平原的过渡地带,东北高、西南低,上游为山区,间有小平原,良口以下逐渐进入丘陵平原区,流域下游地区城镇化率较高,因此从上游到下游其生态状况呈现明显差异。近年来,流溪河沿岸城镇化高速发展,上游河道过度采砂、河道渠道化、过度捕捞、外来物种入侵等问题,使得流溪河流域水生态状况呈明显退化趋势,水功能区达标率低、生物栖息地被破坏、鱼类种类及资源量下降等问题也随之而来[4],因此需要开展全面深入的河流生态健康评估,了解人类干扰程度对流溪河水生态系统的影响。
于2016年6月和12月对流溪河上游至下游20个断面的底栖硅藻和水质进行了调查,分析了流溪河上下游底栖硅藻群落结构和分布特征,明确了影响物种多样性与群落结构的主要水环境因子。通过该项研究,获得了比较完整的流溪河底栖硅藻生物信息,完善了底栖硅藻群落数据库,可为流溪河水质和生态监测提供历史数据和参考。
2 材料与方法
2.1 研究区域及采样点设置
流溪河发源于广州市辖区北部的从化区桂峰山,从东北向西南经从化区、花都区至白云区江村南岗口,纳白坭河、西南涌,于白鹅潭汇入珠江。流溪河流域归属珠江三角洲水系,计至南岗口与白坭河汇流点,干流长为156 km,集水面积为2 300 km2;流域地理坐标为东经113°10′12″~114°2′00″,北纬23°12′30″~23°57′36″,北回归线(N23°26′)横贯流域中部[5]。流域内多年平均气温在21.4℃~21.8℃之间,水资源主要由降雨补给,多年平均降雨量为1 823.6 mm,流域多年平均年径流深为1 226 mm。
在流溪河从上游至下游设置20个采样点,于丰水期(2016年6月)和枯水期(2016年12月),对流溪河底栖硅藻进行2次采样。采样点涉及到的土地利用方式具有多样性,包括山涧溪流、城镇大河、村庄果园、工业园区、市区生活污染、挖山开矿等不同环境状况,从上游至下游,人类活动干扰日益增强。
采样点具体位置如图1所示,各采样点用代码表示(见表1所示)。
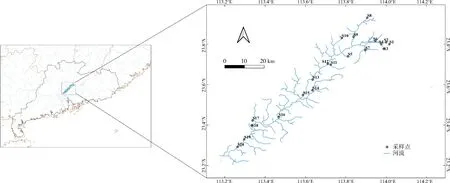
图1 流溪河底栖硅藻采样点分布示意
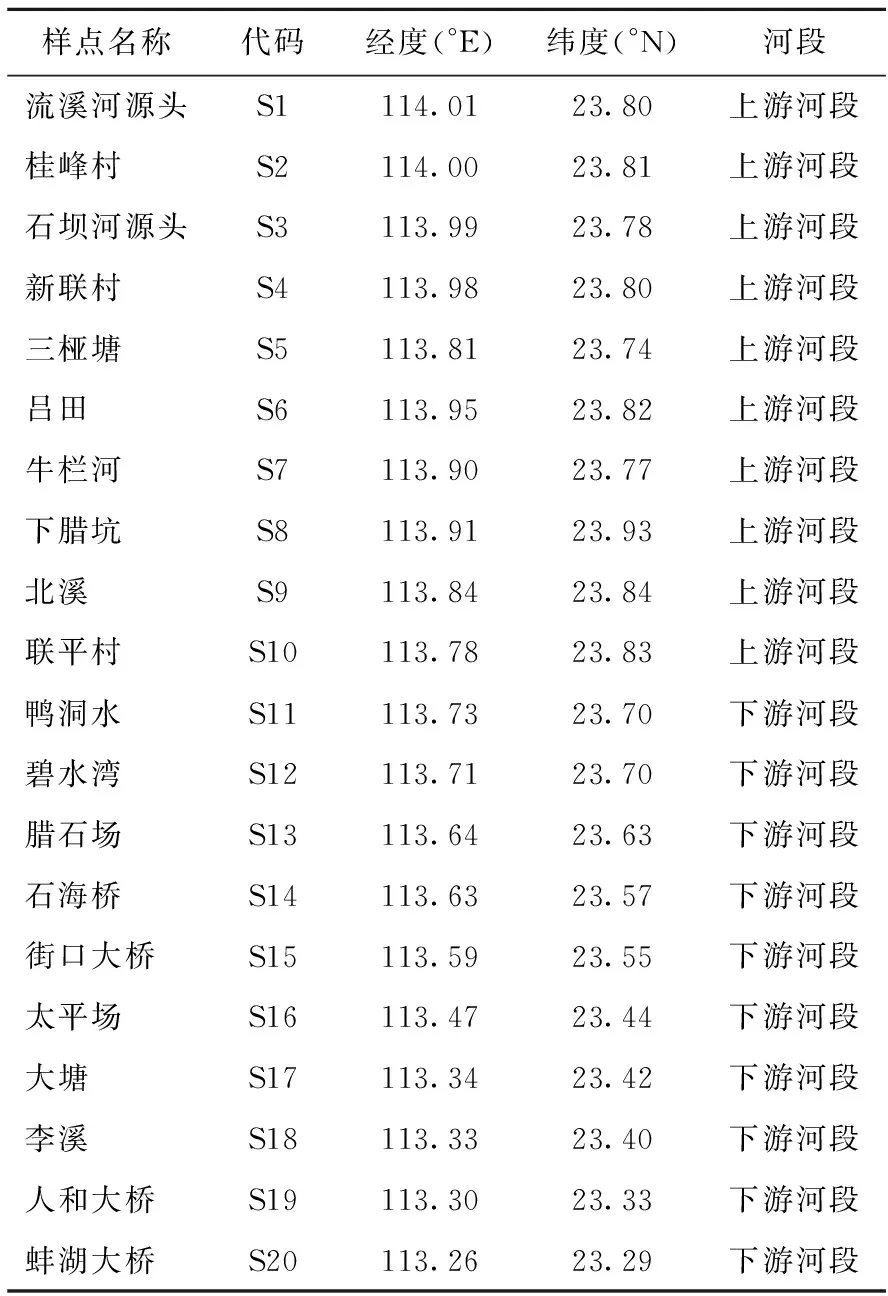
表1 流溪河样点地理坐标、所处河段
2.2 样品采集与处理
2.2.1水环境数据采集与处理
2.2.2底栖硅藻样品采集与处理
采集样品时,尽量选择石头等相似基质进行采样,先用水轻轻地冲去表面异物,再利用牙刷反复刷取附着在基上的底栖硅藻,利用带有100 mL的刻度的透明瓶装取样品,加入甲醛溶液现场固定。样品采样结束后,通过12 h的静置,利用软管除去上清液浓缩至50 mL。取用10 mL样品在高温下(130℃)用浓硝酸氧化处理1 h,冷却后用蒸馏水反复冲洗至中性,用封片胶封片做成永久玻片。最终放大1 000倍的油镜下进行鉴定及统计硅藻细胞数量,每个样品硅藻计数600个。硅藻鉴定参考《珠江水系东江流域底栖硅藻图集》[6]、《中国淡水藻类—系统、分类及生态》[7]。
2.3 数据分析与处理
采用种类数和真辛普森指数表示底栖硅藻物种多样性[8]。真辛普森指数的计算公式为:
(1)
式中:
Pi——物种i在总个体中所占的比例;
S——样本中总的物种数。
使用非参数的Wilcoxon秩和检验,检验两个季节和上下游的主要环境因子(总氮、氨氮、总磷和溶解氧)、物种数和真辛普森多样性指数中位数的差异。以水温、pH值、电导率、溶解氧、浊度、总氮、硝氮、亚硝氮、氨氮、总磷、正磷和可溶性硅作为解释变量,对硅藻分类单元(属)进行RDA分析。对物种多度数据进行Hellinger转换,环境因子进行标准化处理。采用前向选择剔除不显著环境因子,分析环境因子对底栖硅藻群落结构的影响。
所有统计分析在R语言平台上用rstatix、vegan、ade4和adespatial等程序包完成。
3 结果
3.1 流溪河主要环境因子的时空变化
两次调查期间,流溪河总氮浓度在0.54~7.65 mg/L之间,均值为1.85mg/L;氨氮浓度在0.01~4.64 mg/L之间,均值为0.42 mg/L;总磷浓度最低值为0.007 mg/L,最大值为0.335 mg/L,均值为0.065 mg/L;溶解氧浓度最低值为4.77 mg/L,最大值为11.82 mg/L,均值为8.84 mg/L。
Wilcoxon秩和检验表明,总氮、氨氮、总磷和溶解氧浓度存在显著的空间差异(P<0.05),丰水期和枯水期,下游样点总氮、氨氮、总磷和溶解氧浓度的中位数均显著高于上游总氮、氨氮、总磷和溶解氧浓度的中位数(见图2)。丰水期和枯水期,总氮、氨氮、总磷和溶解氧浓度的中位数在季节上无显著差异。
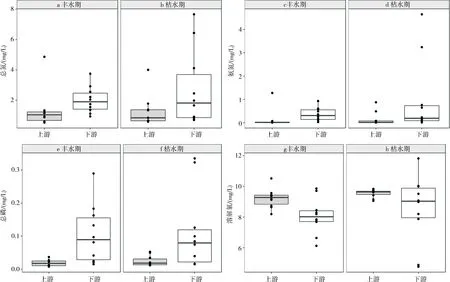
图2 丰水期和枯水期,流溪河主要环境因子的时空变化示意
3.2 流溪河底栖硅藻的多样性的时空变化
丰水期和枯水期的调查中,每个监测站点的硅藻种类数在16~62之间,检出种类最少样点为枯水期的S8(下腊坑),检出种类数最高为丰水期的S10(联平村)。真辛普森多样性指数最小值是丰水期S4(新联村,1.35),最大值为丰水期的S12(碧水湾,18.47)。
非参数分析表明:枯水期,下游种类数、真辛普森多样性指数的中位数显著高于上游种类数、真辛普森多样性指数的均值(P<0.05,见图3);丰水期,底栖硅藻种类数和真辛普森多样性指数空间差异不显著,但下游种类数和真辛普森多样性指数的中位数大于下游种类数和真辛普森多样性指数的中位数(见图3a,c)。枯水期下游的真辛普森多样性指数显著高于丰水期的真辛普森多样性指数(P<0.05),上游的真辛普森多样性指数季节差异不显著(见图3c,d);上游和下游的种类数季节差异均不显著(见图3a,b)。
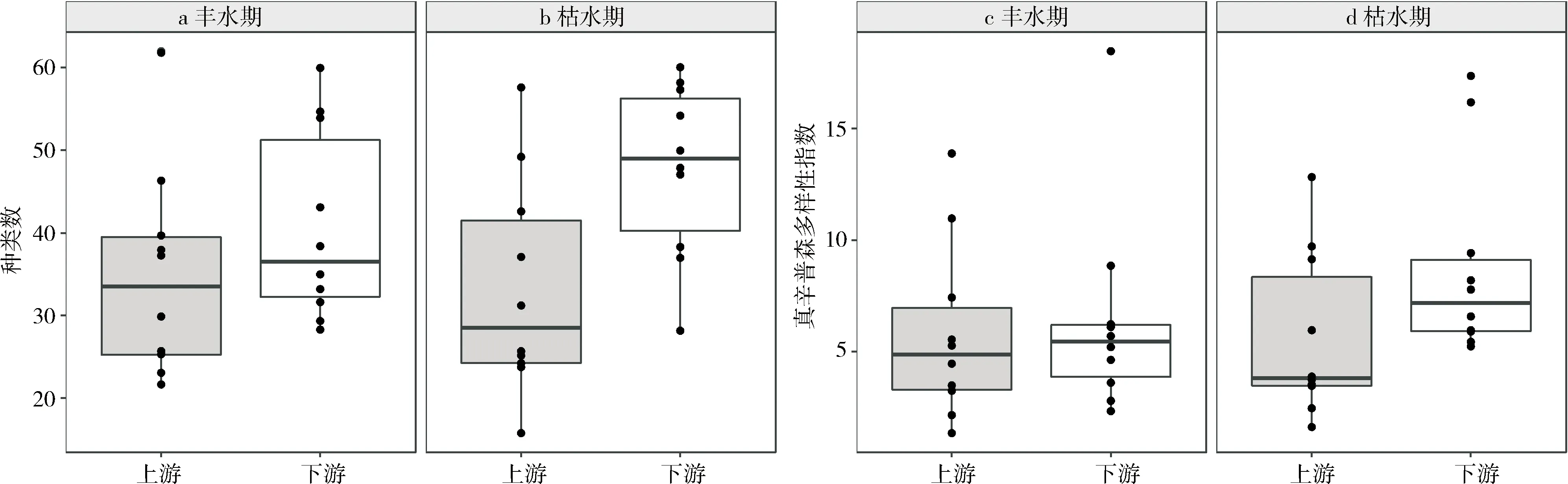
图3 流溪河底栖硅藻多样性的时空变化示意
3.3 流溪河底栖硅藻群落结构
3.3.1种类组成
从两次采集的样品中,共鉴出2纲6目10科54属207种(含变种,见图4)。从纲水平看,以羽纹纲硅藻为主(196种),中心纲硅藻仅11种。从目水平看,双壳缝目种类最多(119种),其次分别是管壳缝目(28种)、单壳缝目(26种)、无壳缝目(13种)、圆筛藻目(11种)以及短壳缝目(10种)。科内的种类多样性分别为舟形藻科(90种)、曲壳藻科(26种)、菱形藻科(23种)、异极藻科(15种)、桥弯藻科(14种)、脆杆藻科(13种)、圆筛藻科(11种)、短壳缝科(10种)、双菱藻科(4种)和窗纹藻科(1种)。
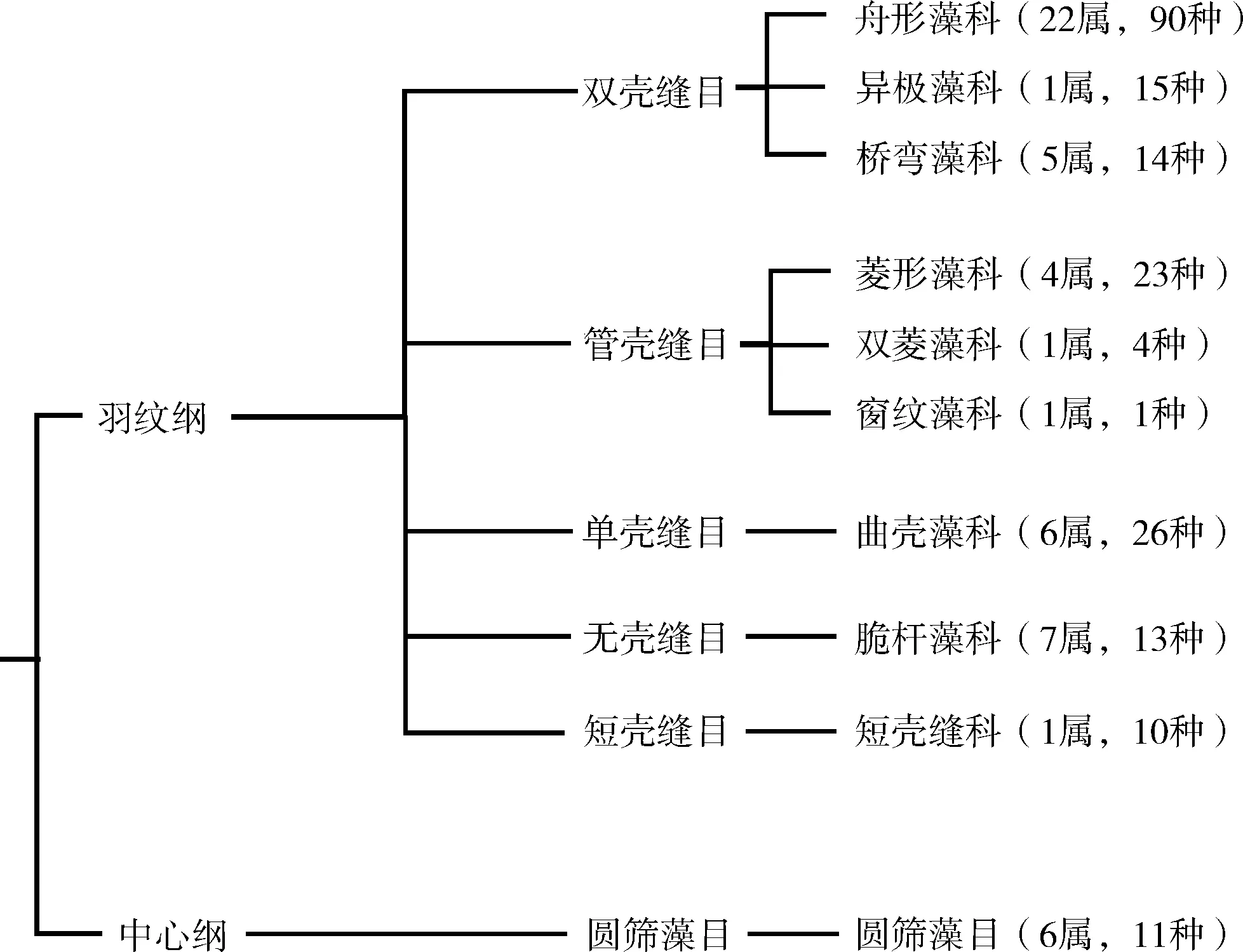
图4 流溪河底栖硅藻分类结构与物种组成示意
丰水期,在20个样点中占据优势的底栖硅藻种类有29种,上游和下游均占据优势的种类有11种,如溪生曲丝藻(Achnanthidium rivulare)、极小曲丝藻(Achnanthidium minutissimum)和微小异极藻(Gomphonema parvulum)等;仅在上游占据优势的种类有8种,如华丽双菱藻(Surirella splendida)、肋缝菱形藻(Nitzschia frustulum)和里德舟形藻(Navicula riediana),上游样点底栖硅藻第一优势种多为溪生曲丝藻(A. rivulare),仅在下游占据优势的种类有10种,如越桔泥生藻(Luticola goeppertiana)、偏肿泥生藻(Luticola ventricosa)和链状曲丝藻(Achnanthidium catenatum)等,下游河段样点第一优势种多为异极藻属和菱形藻属种类,如微小异极藻(G. parvulum)、谷皮菱形藻(Nitzschia palea)(见表2)。
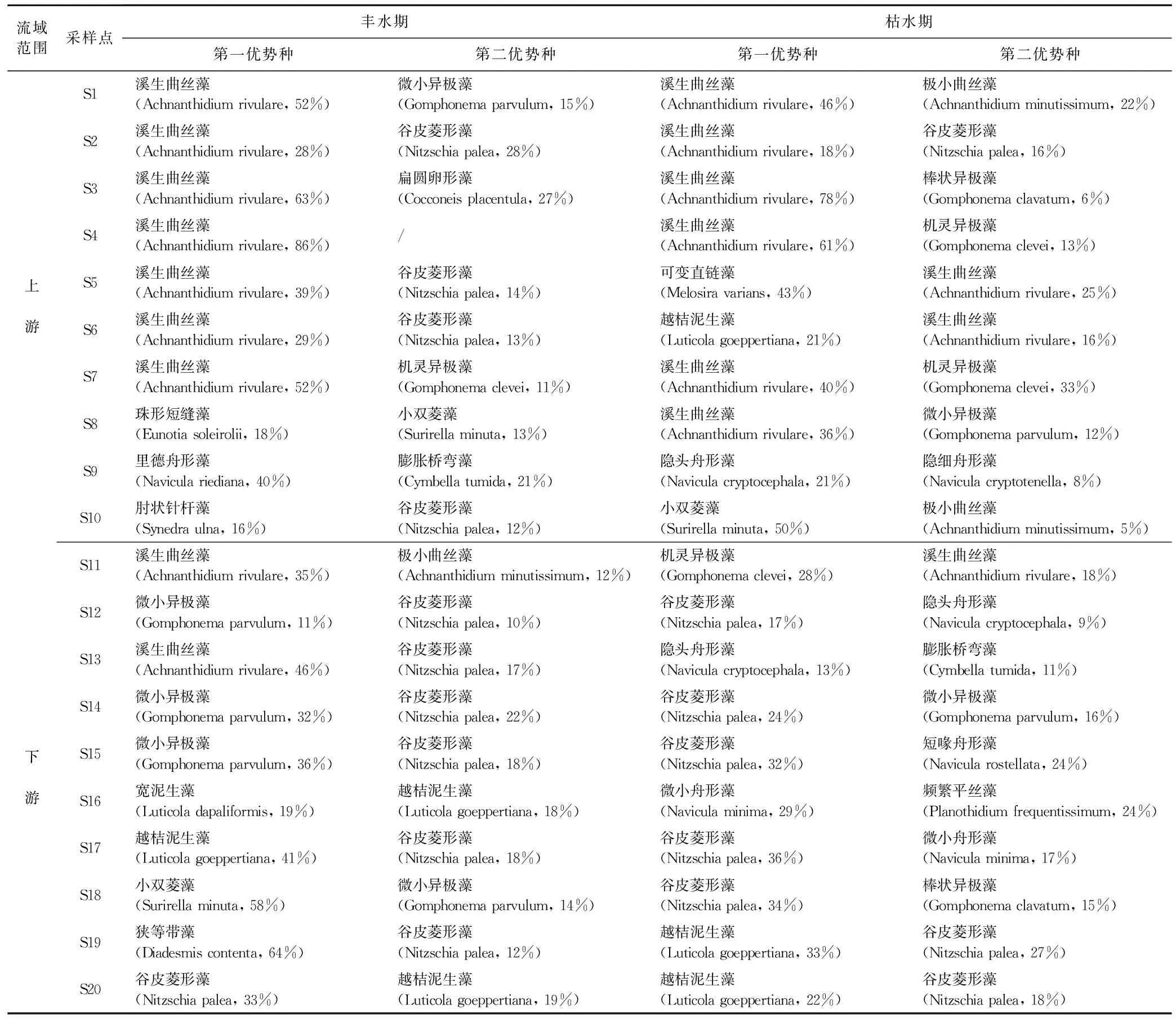
表2 丰水期和枯水期,20个采样点底栖硅藻第一、第二优势物种及其相对丰度
枯水期,优势种种类有32种,上游和下游均占据优势的种类有12种,溪生曲丝藻(A. rivulare)、极小曲丝藻(A. minutissimum)、微小异极藻(G. parvulum)和棒状异极藻(Gomphonema clavatum);仅在上游占据优势的种类有9种,如小双菱藻(Surirella minuta)、小内丝藻(Encyonema minutum)和近赫德森曲壳藻单缝变种(Achnanthes subhudsonis var. kraeuselii)等,小双菱藻在S10样点(联平村)成为第一优势种,越桔泥生藻在S6样点(吕田)成为第一优势种(表2);仅在下游占据优势种类有11种,如微小舟形藻(Navicula minima)、瞳孔鞍形藻(Sellaphora pupula)、频繁平丝藻(Planothidium frequentissimum)和胀大桥弯藻(Cymbella turgidula)等,S19(人和大桥)和S20(蚌湖大桥)以越桔泥生藻(L. goeppertiana)和谷皮菱形藻(N. palea)为第一优势种和第二优势种(见表2)。
3.3.2底栖硅藻群落结构的时空变化
流溪河底栖硅藻主要以曲丝藻属、异极藻属、舟形藻属、菱形藻属、泥生藻属和双菱藻属的种类组成。群落结构存在明显空间异质性。丰水期,上游河段中曲丝藻属种类的相对丰度明显高于下游河段(见图5a);枯水期,上游和下游河段曲丝藻属相对丰度均值差异更加明显(见图5b)。丰水期和枯水期,下游河段舟形藻属、菱形藻属和泥生藻属的相对丰度均明显高于上游河段;异极藻属种类相对丰度均值无明显时空变化。
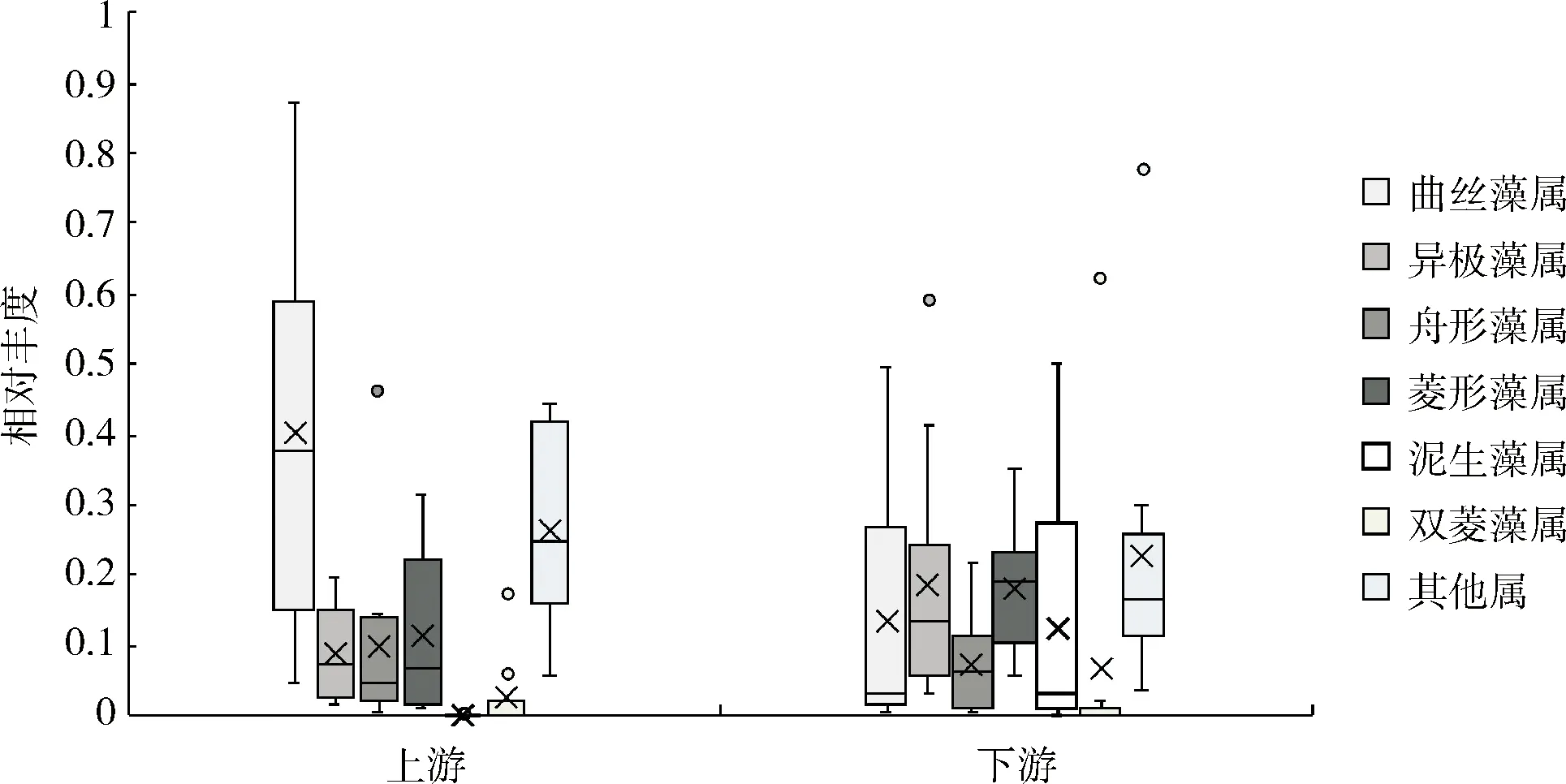
a 丰水期
b 枯水期图5 底栖硅藻群落结构的时空变化示意
3.3.3底栖硅藻群落结构的影响因子
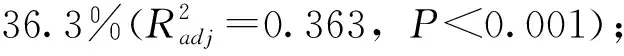

a 丰水期
b 枯水期图6 流溪河底栖硅藻群落的冗余分析示意
2次采样中,上游河段样点和下游河段样点在RDA排序图中能较明显的区分开来,上游河段的样点主要集中在第2、3象限,下游河段样点主要集中在第1、4象限。底栖硅藻分类单元在样点中的分布可知,上游河段曲丝藻属的相对丰度较高,如S1、S3、S4和S7,下游河段中,泥生藻属、菱形藻属和等带藻属的相对丰度较高,如丰水期和枯水期,S19和S20样点泥生藻属的相对丰度较高,S18样点菱形藻属的相对丰度较高。
4 讨论
底栖硅藻作为南亚热带河流的初级生产者,是河流生态系统中基础组成部分[9]。在本研究中,笔者开展了对华南地区典型中小型河流—流溪河的底栖硅藻进行调查,了解流溪河流域底栖硅藻的群落结构以及其环境驱动因素。
通过主要环境因子的时空分布特征,可以很明显的发现,与营养盐有关的环境因子呈现明显的下游高于上游,这明显与人类活动干扰的增加有关,如农业中的氮、磷渗透以及工业废水的排放[10-11]。此外,溶解氧在下游出现下降的趋势可能是因为较高的营养盐和有机质含量会导致细菌的大量繁殖和有机物的分解,进而导致溶解氧浓度降低,甚至出现厌氧状态而威胁水生生物的生长[12-13],本研究中下游溶氧虽然有下降趋势但是整体仍高于溶解氧浓度的临界值(>3 mg/L),说明其环境含氧量满足水生生物的正常生长。
丰水期和枯水期,对流溪河流域底栖硅藻调查,结果显示其优势底栖硅藻十分集中,其中曲丝藻属(Achnanthidium)相对丰度最高可达24.0%,已有研究表明曲丝藻属多生活在河流的源头,适宜流速大、寡营养盐的水体,这与本研究结果相似;而同样占有优势的菱形藻属和异极藻属作为耐污种则更多生活在人类活动干扰较强的下游[14-16]。虽然流溪河物种组成在空间上具有一定差异性,但物种多样性在季节上却并没有呈现明显差异,这说明流溪河流域的生物多样性已经形成了一定的动态稳定性[17],虽然在夏季和秋季会出现一定的增加,但是这可能只是因为径流量增加、温度升高使得水体环境更加适宜藻类的生长与繁殖,而流域整体的生物多样性季节变化并不明显。
综合丰水期和枯水期的硅藻群落结构进一步探讨影响其群落演替的水环境因子,其中,与氮有关的营养盐和海拔无论在丰水期还是在枯水期均有较高的解释率,这与其他学者的研究结果相似[18],这说明对于流溪河流域营养水平是影响硅藻群落结构的最主要因素。在RDA排序图中揭示了一些典型硅藻种类的个体生态属性。其中,溪生曲丝藻(Achnanthidium rivulare)是流溪河上游河段的优势种,这些河段主要以森林用地为主。在丰水期,氨氮、电导率、海拔和pH值和溪生曲丝藻相对丰度呈显著正相关关系,呈显著负相关关系的环境变量为总氮;在枯水期,电导率、海拔、pH值和溶解氧与溪生曲丝藻相对丰度呈显著正相关关系,呈显著负相关关系的环境变量为硝氮、总磷和温度。在下游河段,相对丰度较高的底栖硅藻之一是谷皮菱形藻(Nitzschia palea),这些河段主要以城镇用地为主。在丰水期,与谷皮菱形藻相对丰度呈显著正相关关系的环境变量有总磷和硝氮,呈显著负相关关系的环境变量为海拔;在枯水期,与谷皮菱形藻相对丰度呈显著正相关关系的环境变量主要是正磷。
5 结语
本研究基于对流溪河流域底栖硅藻的水文季节调查,分析了底栖硅藻的群落结构、时空分布特征及其与环境因子的关系。在流溪河底栖硅藻生物信息方面,本次调查提供较为完整的硅藻常见种和指标性物种信息,为建立河流监测需要的底栖硅藻群落数据库夯实了基础,也为流溪河水质监测提供历史数据和参考。通过冗余分析(RDA),发现水质指标尤其是与氮有关的营养盐显著影响影响流溪河中底栖硅藻群落。基于上述分析及实地调查,对流溪河流域生态管理提供3方面的建议:① 持续加强环境监督管理,进一步减少下游工业和生活废水的排入;② 流溪河流域虽然目前已达到生态稳定,但目前两岸人为干扰程度依然很大,因此应该有针对性地开展岸带生态修复工作,改善生物栖息地和生境质量;③ 重视水生生物和连续监测,并基于这些生物数据进行水体健康评价,建立更为科学合理的水生态监测体系。
猜你喜欢
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
小天使·三年级语数英综合(2022年3期)2022-04-17
河南科技(2021年14期)2021-11-28
中国卫生产业(2020年25期)2020-11-30
飞天(2020年9期)2020-09-06
新世纪智能(教师)(2020年2期)2020-05-22
黑龙江水利科技(2020年8期)2020-01-12
作文周刊(中考版)(2017年21期)2017-07-10
故事会(2014年5期)2014-05-14
