
牛磺酸上调基因1在糖尿病及其并发症中的研究进展
2022-12-07 02:09彭俊华江苏大学附属金坛第一人民医院检验科江苏常州213200
现代检验医学杂志 2022年6期
杨 阳,吕 幸,彭俊华(江苏大学附属金坛第一人民医院检验科,江苏常州 213200)
糖尿病(diabetes mellitus,DM)是在遗传因素和环境因素共同作下导致胰岛β-细胞功能异常,临床表现为血糖增高、微血管受损的一种代谢性疾病。多个研究认为非编码RNA( non-coding RNA,ncRNA)在维持β-细胞功能方面起重要作用[1-2],长链非编码RNA (>200个核苷酸, long non-coding RNA,lncRNA)在维持细胞谱系规范中起重要作用[2],胰岛素敏感的lncRNAs调节着胰岛细胞的形成与功能,研究表明1型糖尿病、2型糖尿病(type 2 diabetes mellitus, T2DM)大鼠模型和T2DM病人的胰岛素中均存在着不同的lncRNAs[2-3]。
牛磺酸上调基因1(taurine up-regulated gene 1,TUG1)是最早发现的一个与人类疾病相关的lncRNA,TUG1参与正常细胞增殖、分化及凋亡过程,参与肿瘤的侵袭转移和新生血管生成等过程,参与肿瘤治疗的耐药性、抗辐射性、全身性炎症反应等[4-5],在组织器官的生理性生长发育和机体疾病发生发展中起重要作用。糖尿病与胰岛β-细胞功能异常、慢性炎症、微血管病变相关,本文对TUG1在糖尿病及其并发症中的研究进展进行综述,为糖尿病的预防、糖尿病及其并发症的发病机制研究和治疗提供依据。
1 TUG1概述及功能
YOUNG等[6]于2005年在体外培养发育期小鼠视网膜细胞时加入牛磺酸后发现了一种能被剪接、有多聚腺苷酸尾的 lncRNA,后被命名为TUG1。TUG1长为7.1 kb,定位于人体22号染色体长臂12区2带(22q12.2),含3个转录本[6],在光感受器的形成和视网膜发育过程中起关键作用。
TUG1参与机体的多种生理与病理过程,如调控胰岛素分泌与胰腺β-细胞凋亡、先兆子痫发生、软骨细胞细胞外基质降解、缺氧诱导的心肌损伤等[5,7]。TUG1在多种疾病中过度表达,如多发性硬皮病、慢性肾病、慢性阻塞性肺病等[7]。
多项研究表明TUG1与肿瘤的发生相关。TUG1通过与微小RNA(micro RNA , miRNA)竞争性结合,调节细胞周期依赖性激酶抑制因子的表达,从而影响肿瘤细胞的增殖与凋亡[5]。TUG1在不同肿瘤中显示出不同的作用:一方面作为癌基因,在肿瘤病人体内表达水平升高且预示患者预后差[5,8],可见于膀胱癌、前列腺癌、肾癌、非三阴性乳腺癌、肝细胞癌、胆道及胆管癌、胃癌、结直肠癌、食道癌、胰腺癌、宫颈癌、卵巢癌、肺腺癌、骨肉瘤、甲状腺癌、黑素瘤、急性髓系白血病、肉瘤等;另一方面作为抑癌基因,在肿瘤病人体内表达水平降低[5],可见于非小细胞肺癌、三阴性乳腺癌、神经胶质瘤等。
2 TUG1在糖尿病及其并发症中的基础研究
2.1 TUG1与胰岛β-细胞 糖尿病与机体胰岛素的异常分泌密切相关。胰岛β-细胞的分化、凋亡或功能发生异常,均可导致糖尿病。TUG1表达异常可导致胰岛细胞分泌功能受损。
TUG1与TUG1结合元件(TUG-blinding element, TBE)结合,增加过氧化物酶增殖物激活受体γ (peroxisome proliferator activated receptor γ,PPARγ)共激活因子1α (peroxisome proliferator-activated receptor γ coactivator 1α, PGC-1α)的转录,提高细胞中线粒体的生物能量,调节运动中骨骼肌的肌源性反应[9],抑制胰岛β-细胞凋亡,增加胰岛素的分泌[10],影响糖尿病肾病及慢性肾病的进程[11]。小鼠的心、肝、肾、胰腺、脾和肌肉组织中均可检测TUG1表达,且胰腺中呈高水平表达[7]。下调TUG1表达可诱导小鼠胰岛β-细胞(MIN6)或小鼠胰岛小体中β-细胞的凋亡,抑制胰腺β-细胞的增殖,动物实验与细胞实验结果类似,注入TUG1能降低小鼠空腹血糖和胰岛素耐受水平[12]。
TUG1通过与miRNA结合,调控基因的转录与表达,影响信号通路与蛋白表达,调节细胞的增殖与凋亡。高脂、高糖作用Min6细胞[13]和人正常肠上皮细胞株HIEC-6细胞[14],导致细胞中转录因子特异性蛋白 1 (specificity protein 1, SP1)表达降低,而SP1通过激活miR-188-3p/FGF5轴线途径[13],使乙酰化酶1(sirtuin 1, SIRT1)和磷酸化腺苷酸依赖性活化蛋白激酶(AMP-activated protein kinase,AMPK)表达增加[14],使TUG1表达降低,从而抑制细胞的增殖,促进细胞的凋亡。过表达TUG1通过下调miR-204表达和上调SIRT1/AMPK/乙酰辅酶A羧基酶(acetyl-CoA carboxylase, ACC)信号途径,增加糖尿病小鼠脂肪组织和小鼠脂肪细胞株3T3-L1细胞中的SIRT1,脂肪三酰甘油脂肪酶(adipose triglyceride lipase, ATGL),过氧化物酶增殖物激活受体γ(peroxisome proliferator activated receptor γ,PPARγ)共激活因子1α(PGC-1α)和解偶联蛋白1(uncoupling protein-1, UCP-1)等表达,增加AMPK磷酸化水平,降低糖尿病小鼠中的血糖、胰岛素耐受和脂肪积聚等,促进褐色脂肪修复白色脂肪,从而逆转糖尿病[15]。TUG1还通过对转录后的基因进行修饰来影响疾病的进程,如TUG1调节H3K27甲基化,降低Hes1表达,导致胰岛小体功能紊乱[12]。因此,TUG1在维持胰腺正常功能、调节胰岛素的分泌、促进脂肪细胞的修复等方面具有重要作用。
有研究表明,肥胖诱导T2DM者行部分胃切除术后,体内TUG1和UCP-2表达增加,部分胃切除术通过TUG1/AMPK/ SIRT1/ UCP-2轴线,能缓解高糖或高脂对肠道上皮细胞的损伤[14],进而治疗T2DM和减轻体重。因此,TUG1能调控胰岛β-细胞凋亡,是治疗糖尿病的潜在分子靶标。
2.2 TUG1与糖尿病肾病 糖尿病肾病(diabetic nephropathy,DN)是糖尿病最常见的并发症之一,30%~40%的糖尿病患者会发展为DN。肾小球过滤膜和肾小球系膜是发挥肾小球滤过功能的基本结构。
肾小球上皮细胞,又称足细胞,是肾小球过滤膜的重要组成部分,肾小球足细胞损伤及凋亡,特别是足细胞中线粒体功能紊乱与内质网应急(endoplasmic reticulum stress, ERS)在DN发病机制中起重要作用。高糖环境诱导可促进人近端肾小管足细胞株HK-2细胞[16]、小鼠MPC5细胞[17]的凋亡和ERS[16-17],增加培养细胞中C/EBP同源性蛋白(C/EBP homologous protein,CHOP)的表达,降低PGC-1α和TUG1的表达,而TUG1表达可正向调节PGC-1α水平[18]。高糖处理NRK-52E细胞[19]、原代小鼠足细胞[20]和db/db DN小鼠肾脏组织可使细胞中TUG1表达降低[20-21],且呈剂量-效应关系[19],因高糖环境下碳水化合物反应元素结合蛋白(carbohydrate response element- binding protein,ChREBP)能抑制肾小球足细胞中TUG1的转录[22]。高糖环境还引起肾小球足细胞中线粒体功能发生紊乱,使细胞中活性氧(reactive oxygen species, ROS)水平增加,使TUG1表达降低,敲减TUG1可促进高糖所致的足细胞凋亡和线粒体功能紊乱,反之亦然[19]。DN患者TUG1和PGC-1α表达均降低,但CHOP水平增高,DN小鼠肾小管中TUG1表达水平亦降低[16],TUG1通过调控ERS-CHOP-PGC-1α信号途径,促进肾小球足细胞的凋亡[18]和DN发生。高表达TUG1能激活PGC-1α表达及其转录靶点的表达,改善糖尿病小鼠肾小球中足细胞线粒体中的生物动能[11],TUG1/PGC-1α轴在维护肾小球足细胞线粒体功能稳定中起重要作用[23],影响DN的发生。
lncRNA通过调节miRNA的表达,改变肾小球中足细胞功能和近曲小管的功能[24],减少肾小球足细胞的损伤。TUG1通过与miR-9结合,促进SIRT1表达,导致足细胞线粒体功能紊乱[19];或通过MiR-29c-3p/SIRT1轴线[16],调节足细胞内质网应急[25];或靶向调控miR-34a-5p/ SIRT1信号通路,影响DN中氧化应激的损伤[26];TUG1通过从上述几个方面调控足细胞细胞凋亡,影响高糖环境对肾小球足细胞的损伤。TUG1还通过拮抗miR-377,下调其靶点PPARγ表达,抑制分子内基质如纤溶酶原激活物抑制物1(plasminogen activator inhibitor-1, PAI-1)、转化生长因子β1(transforming growth factor, TGF-β1)、四型胶原、纤维连接蛋白的积聚[27];或通过调节靶点miR-21,促进基底金属蛋白酶组织抑制物-3(tissue inhibitor -3 of matrix metalloproteinases, TIMP3)表达[19],来抑制高糖环境导致肾小球足细胞的纤维化。
肾小球足细胞损伤是导致DN的主要因素,但肾小球系膜发生病变也可影响DN进程。高糖作用下的肾小球系膜细胞中TUG1表达降低,促进糖尿病小鼠出现肾脏增重,出现24h尿蛋白、血尿素氮升高和血肌酐升高等现象。外源性注入TUG1,TUG1通过抑制磷脂酰肌醇3激酶/AKT(PI3K/AKT)信号途径,可减少肾小球系膜细胞细胞外基质的聚集,抑制系膜细胞的增殖和纤维化的形成,从而逆转糖尿病小鼠中尿蛋白和血肌酐升高等表现,延缓DN[21]。
2.3 TUG1与糖尿病视网膜病变 糖尿病视网膜病变(diabetes retinopathy,DR) 是指糖尿病患者的视网膜微血管发生了病变。2010年全球34.6%糖尿病患者中发生了DR,我国>45岁人群中有18.45%糖尿病患者发生了DR。视网膜中的细胞或神经元凋亡、白细胞淤滞、内皮细胞的增殖等因素引起视网膜中神经发生退行性变、血管的通透性增加、新生血管的形成等,最终可发生DR。人视网膜微血管内皮细胞(human retinal microvascular endothelial cells, hRMECs)形态呈树枝状以支持视网膜神经,既供给视网膜的营养需求,又形成屏障保护视网膜。高糖环境可提高hRMECs的细胞增殖活力、细胞迁袭能力和小管生成能力,使hRMECs中TUG1表达增加[28],敲减TUG1可调节miR-145/血管内皮生长因子α(vascular endothelial growth factor-α,VEGF-α)轴线,进而抑制高糖环境下hRMECs的细胞增殖、迁移和小管生成能力,延缓DR的进展[28]。
糖尿病还可导致神经损伤,早期及时的干预能减少氧化应激所致神经损伤。绿原酸,一种来源于咖啡、茶、水果和蔬菜中的生物活性物质,通过TUG1/核因子相关因子2通路,能提高视网膜神经节细胞的活力,减少活性氧聚集,抑制视网膜神经节细胞凋亡,从而使视网膜神经节细胞免受氧化应激的损伤[29]。临床常用局麻药布比卡因可使高糖环境下小鼠背根神经节(dorsal root ganglion,DRG)的活性氧增加,自噬激活,TUG1表达增加[30]。因此,TUG1位点是糖尿病神经损伤的潜在治疗靶点,也可能是研究其它药物治疗糖尿病的靶点。
2.4 TUG1与糖尿病心脑血管病变 流行病学研究认为冠心病(coronary artery disease,CAD)与代谢性疾病如糖尿病密切相关。高糖环境下培养冠心病患者组织及内皮细胞,发现培养组织或细胞中TUG1表达和肿瘤坏死因子α (tumor necrosis factor α,TNF-α)水平均升高,高表达TUG1可促进人脐静脉内皮细胞(human umbilical vein endothelial cells, HUVECs)的细胞增殖、细胞迁移和细胞周期加速,使β-catenin和c-Myc蛋白表达增加,从而促进糖尿病血管硬化的发生和发展,而Wnt信号通路抑制剂如XAV-939则可抑制TUG1对HUVECs的促增殖和促迁移等作用[31]。糖尿病心肌病(diabetic cardio-myopathy, DCM)是指糖尿病患者的心肌结构和功能发生异常,糖尿病患者出现心脏肥大和心脏功能紊乱等,严重时可导致糖尿病患者死亡。在DCM性db/db小鼠心脏中TUG1表达增加,抑制TUG1表达能改善DCM性db/db小鼠心脏舒张期紊乱,敲减TUG1可上调心肌细胞中miR-499-5p表达,能减少小鼠心肌纤维化区域,进而缓解DCM诱导的心脏肥大和舒张期紊乱等[32]。慢性缺氧条件下小鼠心脏成纤维细胞中TUG1表达水平增高[33],通过miR-29c影响心脏的纤维化。因此,TUG1位点可能成为糖尿病心血管疾病的治疗靶点。
2.5 TUG1与糖尿病足综合症 糖尿病足综合症(diabetic foot syndrome, DFS)是一个常见的糖尿病并发症,其病因可能与干细胞,特别是内皮祖细胞(endothelial progenitor cells,EPCs)的功能紊乱有关。骨髓来源EPCs在特定情况如缺氧时能促进细胞增殖、迁移和分化,促进血管形成和缺血后新生血管的生成。细胞实验表明高糖环境能抑制EPCs的迁移、侵袭和小管形成能力,高表达TUG1可逆转EPCs的上述改变。向糖尿病缺血性肢体病小鼠注入TUG1,可调节miR-29c-3p/PDGF-BB/Wnt信号途径,促进EPCs的增殖、迁移和小管生成,加速病侧血管的修复[34]。因此,TUG1参与了DFS及糖尿病缺血性肢体病侧血管修复。
总之,除了高糖、高脂的因素外,药物、食物或手术作用于DM也会影响TUG1表达。TUG1通过影响PGC1表达的改变和miRNA的激活或拮抗作用来调控胰岛素的分泌与胰腺β-细胞的凋亡,另外,TUG1还通过影响细胞间信号传导途径的改变、基因甲基化等因素,最终影响糖尿病及其并发症的发生、发展和治疗,见图1。
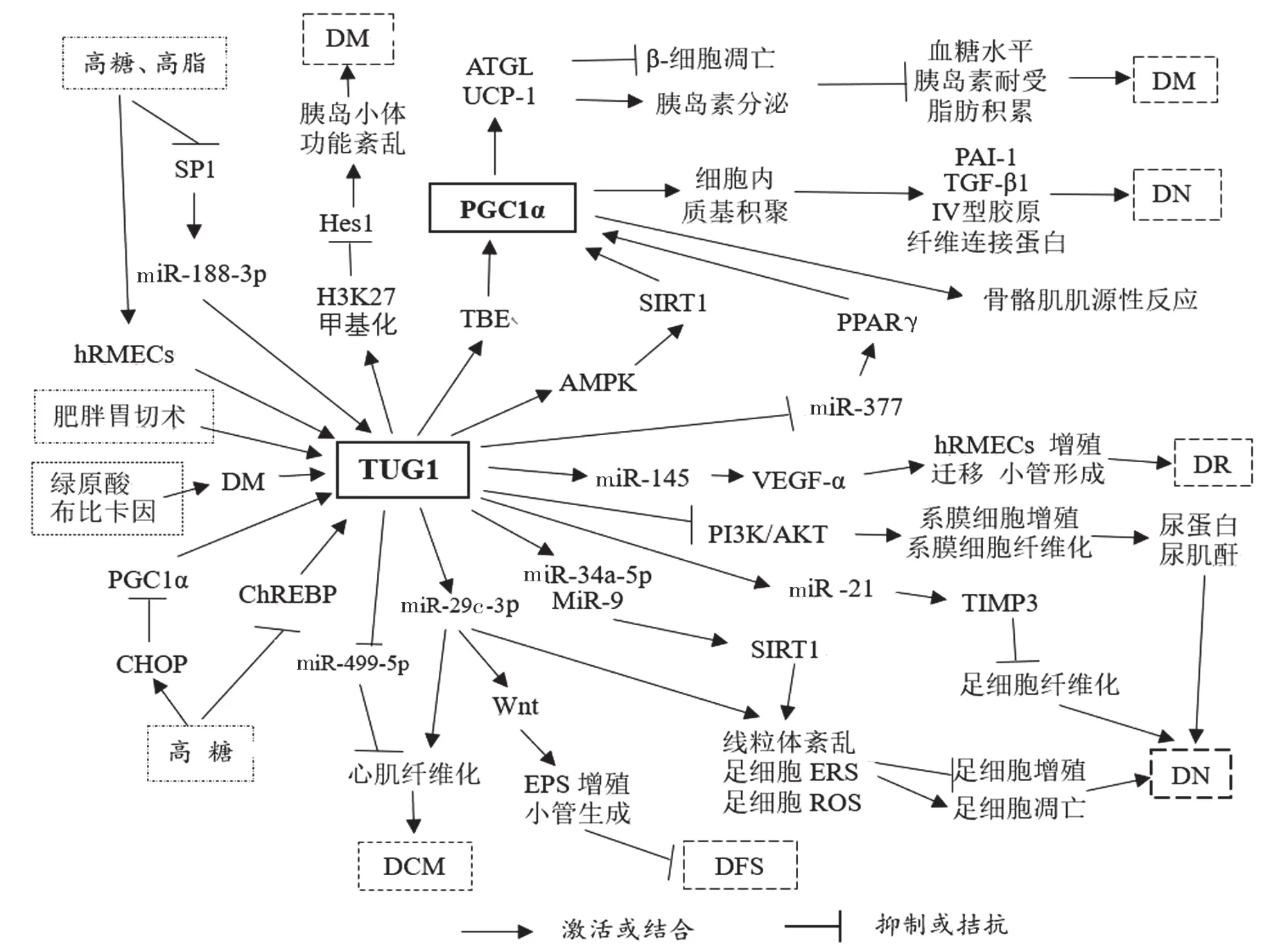
图1 TUG1与糖尿病及并发症的关系
3 TUG1在糖尿病及其并发症的临床研究
TUG1与糖尿病及其并发症间的研究多数为体外的细胞学研究及动物研究,临床研究主要集中在外周血TUG1表达水平和TUG1基因型与DM及其并发症的发病机制及诊断方面。多个研究采用实时荧光定量PCR(qRT-PCR),结合内参用2-ΔΔCt法计算TUG1的相对表达量[35-39],结果表明DM患者外周血[35]、外周血白细胞[36]、外周血单个核细胞[37]中TUG1表达水平均明显高于健康对照组,且T2DM并发睡眠不足者或早期DN者外周血中TUG1水平明显高于单纯T2DM组[36]。糖尿病缺血性中风患者外周血清TUG1和miR-9表达水平明显升高,miR-106a表达降低[38]。TUG1高水平表达可促进T2DM和早期DN发生[39],且与T2DM患者空腹血糖、糖化血红蛋白和病程等呈正相关[36],因此,外周血TUG1表达水平对诊断T2DM有较高的灵敏度和特异度[35]。
TUG1基因型不同也可导致DR发病机率及治疗用药的差异。MOHAMMAD等[40]用PCR法进行基因多态性分析,结果显示携带TUG1A/G和MIAT T/C或C/C基因型容易发展为DR,携带TUG1 A或MIAT T/C的患者对血管内皮生长因子抑制剂阿帕西普的治疗效果较差,见表1。
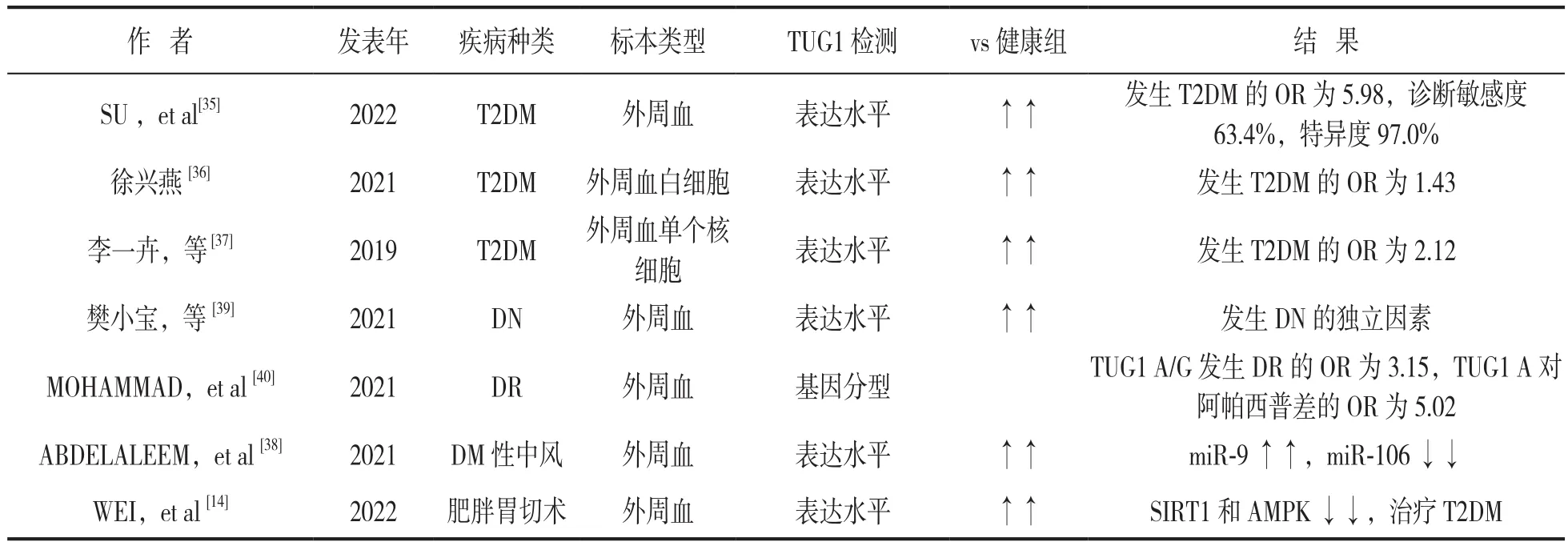
表1 TUG1检测在糖尿病及其并发症诊断及预后的研究情况
总之,外周血TUG1高表达是DM或DN发生的独立危险因素,TUG1可能成为DM及其并发症诊断的标志物。虽然TUG1作为一个lncRNA可能参与机体所有组织和细胞的生理过程,对某一单个疾病的诊断、治疗及其预后判断的敏感度较差,但随着研究的不断深入,TUG1有可能成为DM及其并发症的诊断、治疗及预后判断等的一个辅助因子。
4 小结与展望
TUG1是糖尿病研究中的一个热点问题,但目前尚存在一些争议问题,诸如DN小鼠肾小球或肾小管、DM胰腺中TUG1表达降低,高糖环境下hRMECs细胞或冠心病患者的内皮细胞中TUG1表达增加,且DCM性db/db小鼠心脏中的TUG1表达也增加,这些不同的实验结果是否与不同细胞的特异度相关,是否与实验研究模型、人种、动物或细胞种类以及TUG1检测所用引物、体内实验与体外实验控制条件相关,尚需要进一步的研究证实。多个研究表明T2DM或DN患者外周血中TUG1升高,这种升高是因为胰腺中TUG1表达降低而引发的代偿性升高,还是因为细胞与细胞外液中的分配不同,也需要进一步的研究。
未来,在进一步明确TUG1与DM及其并发症的作用机理的基础上,研发出能调控TUG1与靶点结合的靶基因药物,或以此靶点来研究药物的作用机理,从而更好、更精准地治疗和缓解糖尿病及其并发症。
猜你喜欢
医学信息(2022年9期)2022-11-27
天津医科大学学报(2021年4期)2021-08-21
中成药(2018年6期)2018-07-11
中成药(2017年8期)2017-11-22
中外医疗(2016年15期)2016-12-01
中成药(2016年8期)2016-05-17
中国病理生理杂志(2015年8期)2015-12-21
医学研究杂志(2015年9期)2015-07-01
中国药理学通报(2014年2期)2014-05-09
中国医学科学院学报(2014年6期)2014-03-11
