
非生长季放牧对黄河上游草甸种子产量的影响
2022-12-02 05:47:46陈安群张露娜柴诗杰王文瑞刘银占韩士杰苗原
河南师范大学学报(自然科学版) 2022年6期
陈安群,张露娜,柴诗杰,王文瑞,刘银占,韩士杰,苗原
(河南大学 生命科学学院,河南 开封 475004)
草地生态系统约占全球陆地面积的40%,对陆地生态系统应对气候变化有着重要影响.此外,草地作为重要的畜牧业生产基地,为人类提供大量的肉类和奶制品,具有十分重要的经济价值[1].放牧是草地生态系统重要的土地利用方式之一.在放牧过程中,动物通过采食、踩踏和粪便排泄等方式作用于植物生长与繁殖[2],影响着生态系统物种组成与结构[3].首先,动物通过采食等可直接改变植物光合产物的积累,影响生物量[4],进而影响到植物光合产物向花、果实、种子等生殖器官的分配[5-7].例如,放牧情况下内蒙古荒漠地区重要的建群种垫状锦鸡儿(Caraganatibetica)的果实数显著减少[6].放牧对天然草原上羊草(Leymuschinensis)的有性生殖产生制约,影响羊草的自然更新[8].其次,动物还可通过践踏和粪便等改变土壤结构,以及土壤中水分和养分含量,间接影响植物生物量积累以及生殖分配.
与此同时,植物可以主动调整其生长和繁殖策略应对放牧对其造成的影响.种子产量作为植物重要的繁殖指标,是土壤种子库的直接影响因素,关系着草原植物新生苗补充以及草原的抗干扰能力[9-10].在受干扰情况下,植物可以通过调节种子大小和种子数量之间的权衡关系,最大化其生存适应性.但因不同植物的生长特性、适口性和生活史对策存在差异,种子大小和种子数量之间的权衡关系千差万别,因此种子产量对放牧干扰的响应也不尽相同.有研究发现,放牧减弱三叶草种间竞争,导致其生产的小种子数量较多;而鸭茅因其花穗高易被采食,成穗率低,在种子生产时采取少而大的策略来应对放牧干扰[11].
尽管关于放牧对种子产量的影响机制的研究很多,但这些研究多关注于生长季放牧对其造成的影响,关于非生长季放牧对种子产量影响的研究较少.非生长季与生长季植物生长的状态不同,这两个季节放牧对植物的影响机制可能也存在差异.若通过生长季放牧的实验结果推测非生长季放牧对植物生长和繁殖(如种子产量)的影响可能存在一定的偏差.为更准确了解非生长季放牧对植物生长繁殖的影响,本研究在青藏高原的高寒草甸选取非生长季放牧和对照区域,探讨非生长季放牧对草地生态系统种子产量的影响,并探索其可能的影响机制.
1 材料与方法
1.1 研究地点
研究地位于四川省阿坝藏族羌族自治州红原县境内(32°48′N,102°33′E,海拔3 500 m).该地属于大陆性高原温带季风气候,年均温1.4 ℃,最高和最低气温分别为10.9 ℃和-10.3 ℃.非生长季时间较长,大约有158 d.年均降雨量为749 mm,大多集中在5至10月份.植被类型为高寒草甸.植物种类大约有23种,植被覆盖率70%~90%.实验地点非生长季放牧强度为3~5头牦牛/hm2.非生长季放牧时间为10月中旬至次年5月初.
1.2 种子采集方法
采用成组比较的实验设计,在2007年开始设置的围封对照区和自20世纪90年代末开始一直进行处理的非生长季放牧区选取5对区域.每对区域内的两种处理下分别选取10个物种,分别为:矮地榆(Sanguisorbafiliformis)、发草(Deschampsiacespitosa)、美丽龙胆(Gentianaformosa)、花葶驴蹄草(Calthascaposa)、条叶银莲花(Anemonetrullifoliavar.linearis)、高山紫菀(Asteralpinus)、高原毛茛(Ranunculustanguticus)、华扁穗草(Blysmussinocompressus)、矮泽芹(Chamaesiumparadoxum)和川西北苔草(Carexenervis).每对区域内每个物种标记2株,共标记10株,然后记录每株植物的开花数和果实数.果实成熟时收集所有果实中的种子,并统计每株植物上的饱满种子数量和总的种子数量.收集到的种子65 ℃烘干48 h后称重,得到每株植物的种子产量.通过以上得到的数据,参照以前的研究[5-7],分别计算种子大小、坐果率和结实力.
1.3 数据分析
首先将植物物种和非生长季放牧作为两个实验因素,对种子产量(每株植物的种子质量)、种子数量、种子大小、花朵数、坐果率和结实力分别进行双因素方差分析.不同物种间的多重比较采用Duncan检验.之后每个单独的物种进行单因素方差分析,检验非生长季放牧对每种植物种子产量及相关指标的影响.最后以种子产量以及放牧引起的种子产量变化作为因变量,与种子数量、种子大小、花朵数、坐果率和结实力以及上述指标对放牧的响应分别进行简单线性回归和多元逐步回归分析.所有统计分析通过SPSS 26.0进行.
2 结 果
2.1 种子产量
物种的不同和放牧对种子质量均有显著影响(P<0.001,表1).不同物种种子产量由大到小依次为:矮泽芹,发草,高山紫菀,高原毛茛,条叶银莲花,矮地榆,美丽龙胆,川西北苔草,华扁穗草,花葶驴蹄草(表1).综合10个物种来说,放牧使得种子产量降低28%(表1,表2).由于放牧与物种之间存在显著的交互作用(P<0.01,表2),故不同物种对放牧的响应不同,放牧显著增加高原毛茛的种子产量(表1,附表Ⅰ),降低矮地榆、发草、美丽龙胆、花葶驴蹄草、条叶银莲花和高山紫菀的种子产量,降低量分别为32,374,42,17,116,147 mg(所有的P<0.05,表1,附表Ⅰ).
2.2 种子数量
对于种子数量来说,物种之间亦存在显著差异(P<0.001,表1,表2).综合来看,非生长季放牧使得种子数量降低22%(P<0.001,表1,表2).物种与放牧之间存在显著的交互作用(P<0.001,表2).对于单个物种来说,非生长季放牧显著增加高原毛茛的种子数量(+420),显著降低发草、美丽龙胆、花葶驴蹄草、条叶银莲花和高山紫菀的种子数量(P<0.05),对矮地榆种子数量的减少达到边际显著水平(P<0.10),对华扁穗草、矮泽芹和川西北苔草的种子数量无影响(表1,附表Ⅰ).
2.3 种子大小
物种与非生长季放牧对种子大小均有显著影响(P<0.01表2).不同物种间,条叶银莲花的种子最重,为2.20 mg,显著高于其他物种.不同物种种子大小对非生长季放牧的响应不同,非生长季放牧显著增加川西北苔草的种子质量(+0.09 mg),显著降低高原毛茛(-0.07 mg)、高山紫菀(-0.14 mg)的种子质量,对于花葶驴蹄草(-0.05 mg)、条叶银莲花(-0.31 mg)的种子质量的降低仅达到边际显著水平(P<0.10).但总的来说,放牧使得质量大小降低9%.
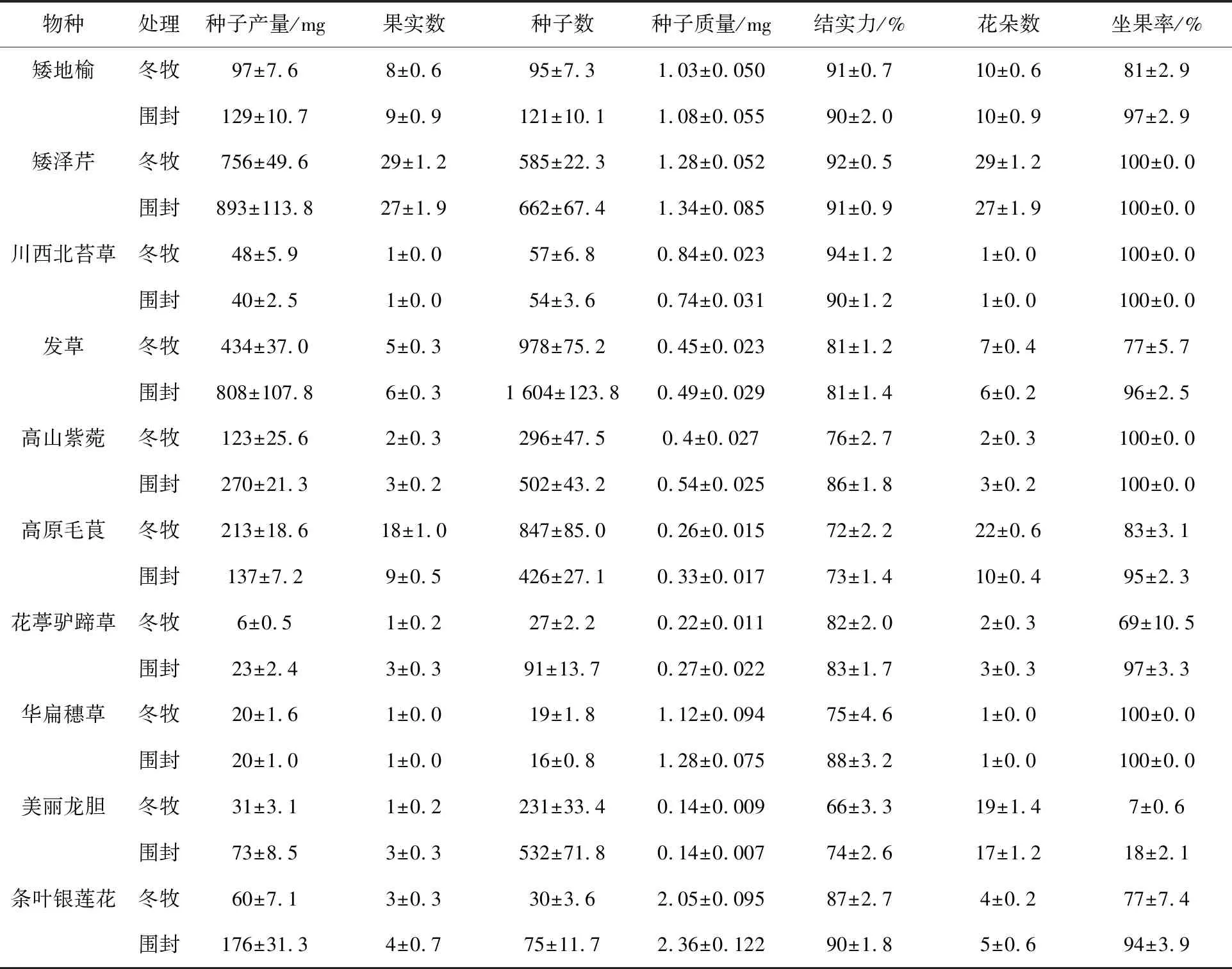
表1 不同处理下高寒草甸植物种子产量、果实数、种子数、种子质量、结实力、花朵数、坐果率(平均值±标准误差)

表2 放牧与不同物种对种子产量、种子数、种子质量、花朵数、坐果率和结实力影响的两因素方差分析结果
2.4 花朵数
不同物种间花朵数差异显著(P<0.001,表1,表2),多重比较的结果显示,除条叶银莲花和花葶驴蹄草之间、花葶驴蹄草和高山紫菀之间以及高山紫菀、华扁穗草和川西北苔草之间的差异性不显著以外, 其他物种间差异均达到显著水平.综合10个物种,非生长季放牧使得花朵数显著增加21%.对于单个物种来讲,发草和高原毛茛的花朵数在放牧的情况下得到显著提高,绝对增加量分别为1.30和12.1,而花葶驴蹄草和高山紫菀的花朵数却显著降低,降低量分别为1.10和0.86,其他物种的花朵数对非生长季放牧没有显著响应(表1,附表Ⅰ).
2.5 坐果率
不同物种和非生长季放牧对坐果率的影响均达到显著水平(P<0.001,表1,表2),并且两者之间存在显著的交互作用.美丽龙胆的坐果率最低,为13%,显著低于其他9个物种;川西北苔草、高山紫菀、华扁穗草和矮泽芹之间差异不显著;高原毛茛、矮地榆、发草、条叶银莲花和花葶驴蹄草之间差异不显著(表1).综合10个物种来讲,放牧处理下坐果率降低10%.分物种来看,放牧显著降低矮地榆、发草、华扁穗草、美丽龙胆、花葶驴蹄草和高原毛茛的坐果率,对条叶银莲花坐果率的降低达到边际显著水平(P=0.06),对其他物种的坐果率影响不显著.
2.6 结实力
不同物种间结实力差异显著(P<0.001,表2),由大到小依次为:川西北苔草,矮泽芹,矮地榆,条叶银莲花,花葶驴蹄草,华扁穗草,高山紫菀,发草,高原毛茛,美丽龙胆(表1).放牧显著降低结实力3%(P=0.003,表1),由于物种与非生长季放牧之间交互作用的存在,不同物种的结实力对非生长季放牧的响应不尽相同.放牧使得川西北苔草结实力增加3%(P=0.09),却显著降低华扁穗草和高山紫菀的结实力,分别降低13%和10%,对美丽龙胆的降低仅达到边际显著水平,对其他物种无影响(表1,附表Ⅰ).
2.7 种子产量变化与其余指标变化的关系
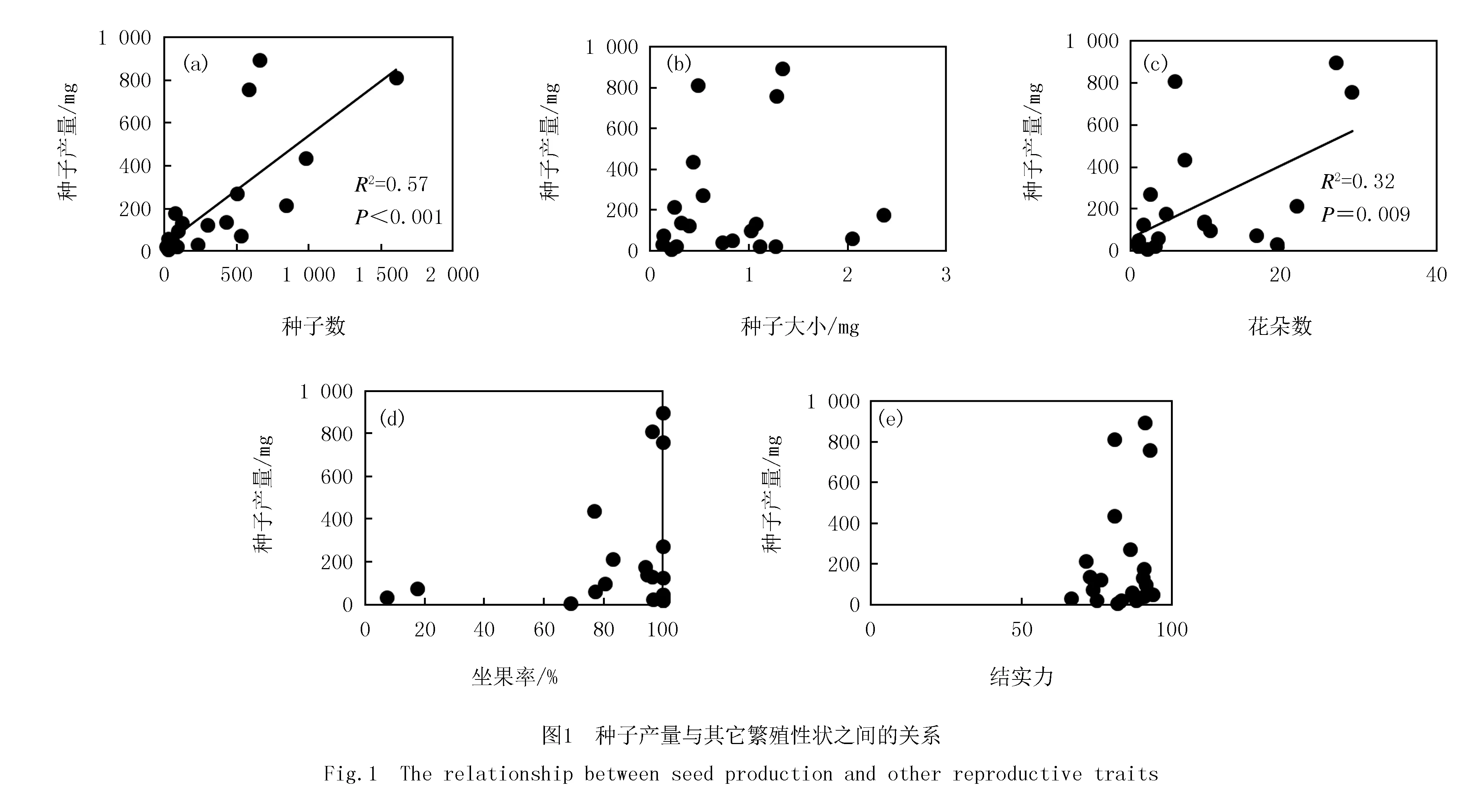
种子产量与其他指标之间的简单线性回归分析表明,种子产量与种子数量、以及花朵数之间存在显著的正相关关系,其R2值别为0.57和0.32,其余指标与种子产量和花朵数之间的关系并不显著(图1).利用多元线性逐步回归分析种子产量与其他5个指标的关系发现,种子数和花朵数可共同解释种子产量变化的66%(P<0.001).并且根据标准化系数来看,种子数量的贡献最大(0.63),其次是花朵数(0.33).利用放牧对不同指标的处理效应进行回归分析则发现,放牧引起的种子数量的变化可以解释种子产量变化的73%,其余指标的变化均与放牧引起的种子产量无显著关系(图2).
3 讨 论
3.1 非生长季放牧对种子产量的影响
综合所选的10个物种,非生长季放牧显著降低种子产量(表1,表2).相关分析的结果显示,种子产量的变化主要与种子数量的变化密切相关(图1,图2).与本结果类似,杜利霞等[12]在内蒙古草原的研究发现,放牧降低冷蒿种子产量,并且其种子产量与花序数和成熟种子数显著相关.花作为植物重要的繁殖器官,其数量关系着植物的有性繁殖能力.植物在秋季形成花原基或花序原基,经过冷冻和春化,在适宜的光照和温度下分化形成花芽.尽管有的研究认为,生长季结束后收割植物地上部分或放牧等土地利用方式可以为植物提供更多的生长空间,还可以增加光的通透性[13],进而促进生殖枝的分化生长,从而提高种子产量[14].但非生长季放牧过程中,牲畜可能通过采食、践踏等方式破坏秋季形成的花原基或花序原基,导致来年生长季分化形成的花芽数降低[5],进而降低种子产量[15].其次,该研究中非生长季放牧时间一直持续到5月中旬,此时很多植物开始返青.由于非生长季放牧去除了植物地上部分以及部分凋落物,植物群落可以得到更多的光照,提前其返青期[14].在非生长季放牧的晚期,部分植物开始萌发分蘖.新长出的植物部分,对于牲畜来说适口性更好[16],这会造成大量植物新生部分被采食.最终非生长季放牧处理下植物分蘖数和花芽数减少,花朵数降低[7],导致种子产量下降.此外,即使有些物种花朵数未受非生长季放牧影响,但由于坐果率显著降低(表2,图1)[6],使得在同样的花朵数甚至更多的花朵数的情况下,植物种子数量减少[14],造成种子产量降低,如本研究中的矮地榆、发草、美丽龙胆和条叶银莲花.这些结果表明,非生长季放牧的效应会持续到来年生长季,影响植物的繁殖策略,如减少种子产量.但植物也可以通过增加无性繁殖部分的投入来应对放牧影响,达到该生长条件下最优的生存适合度[11].因此关于植物如何调整其繁殖分配响应非生长季放牧的影响还需进一步研究.

3.2 不同物种种子产量对非生长季放牧的响应
尽管10个物种的综合结果显示非生长季放牧降低种子产量,但是由于不同植物物种生物和生态学特性的差异,对放牧干扰的响应也千差万别.在本研究中放牧显著降低矮地榆、发草、美丽龙胆、花葶驴蹄草、条叶银莲花和高山紫菀的种子产量,却显著提高了高原毛茛的种子产量,对华扁穗草、矮泽芹和川西北苔草的种子产量无显著影响.之前在人工建植草地的研究中也发现,白三叶草由于其植株矮小,再生能力强,在放牧干扰后依然保持较高的生殖生长;但红三叶草植株较高,易被牲畜采食,生殖枝数量在放牧条件下明显降低[11].另外,物种返青期的差异也会造成对非生长季放牧的不同响应[16].
种子产量是种子数量和种子大小的综合体现.足够的种子数量可保证植物种子得以广泛传播,还可大量补充土壤种子库,为幼苗的萌发以及生态系统的演替提供保障.而种子作为遗传物质的载体,它的大小关系到种子中遗传物质的质量,以及种子在土壤中的存留时间和对不利环境的承受力[17],是种子活性的重要指示指标[18].一般认为小种子更易散播;而大种子比小种子更易存活,萌发率更高.在受到干扰(如放牧)情况下,植物可以权衡种子数量和种子大小之间的关系[11],最大化自身的生存利益,因而形成不同物种种子产量对放牧干扰的不同响应.放牧情况下一般会有以下几种情况:1)种子数量和种子大小对非生长季放牧均无显著响应,种子产量也无明显变化,如华扁穗草和矮泽芹.2)种子数量和种子大小都减少,种子产量自然降低,如本研究中花葶驴蹄草、条叶银莲花和高山紫菀种子产量对放牧的响应.3)种子数量和种子大小两者中一方减少,另一方未受影响;结合本研究中相关分析的结果(图1,图2),此时种子质量的变化一般会与种子数量的变化保持一致.如矮地榆、发草和美丽龙胆的种子数量降低,种子大小未改变,它们的种子质量也降低.而川西北苔草的种子大小显著增加,种子数量未改变,它的种子质量也没有显著变化.4)种子数量增加而种子大小减少,由于种子产量和种子数量之间密切的相关关系(图2),这种情况下放牧提高种子产量,如本研究中的高原毛茛.非生长季放牧降低大部分物种的种子产量,可能这些物种将更多的资源用于无性繁殖,从而可以更好、更快地占据更多的空间和光照[9,15].而高原毛茛采取生产大量的小种子,使得种子在成熟之后可以快速扩散,增大其分布范围,大量补充土壤种子库,在来年种子萌发时可以占据因放牧造成的空位.还有一些物种种子产量对非生长季放牧无显著影响,可能是这些物种在生长早期放牧干扰的刺激下再次分蘖,产生补偿效应[6].总之,植物根据其自身的生物学和生态学特性调整各自的繁殖对策,最终目的都是增强其在干扰环境下的适应能力.
综上所述,本实验选取黄河上游青藏高原草甸草原10种植物,研究非生长季放牧对种子产量及其他生殖指标的影响.实验结果表明,放牧显著降低大部分物种的种子产量,但增加高原毛茛的种子产量,这可能与不同物种的生物学和生态学特性差异,以及不同物种在面对放牧干扰下的繁殖对策选择有关.并且在该实验地点,种子产量与种子数量和花朵数显著相关.因此在植物返青时,应及时停止非生长季放牧,以保护植物新萌发的分蘖,保证植物具有足够的生殖枝数量,进而获得充足的花朵数和种子数量.非生长季放牧对有性生殖指标影响的研究可以帮助我们更好地理解不同时期放牧对植物生长繁殖的不同作用.研究非生长季放牧条件下种子产量的响应,有助于了解土壤种子库的变化[19],以及植物群落的更新演替,为草原生态系统的管理提供数据支持.
附 录
附图见电子版(DOI:10.16366/j.cnki.1000-2367.2022.06.007).
猜你喜欢
云南化工(2021年9期)2021-12-21 07:43:46
中老年保健(2021年11期)2021-08-22 03:15:20
今日农业(2021年8期)2021-07-28 05:56:04
阅读(低年级)(2020年9期)2020-11-06 06:23:05
文苑(2020年6期)2020-06-22 08:41:46
河北果树(2020年2期)2020-01-09 11:15:07
今日农业(2019年11期)2019-08-15 00:56:32
今日农业(2019年11期)2019-08-15 00:56:32
家庭医学(2018年4期)2018-07-18 04:31:58
散文诗(2017年17期)2017-08-15 00:47:32
