
山东泰山地区猕猴桃品种间抗寒性调查与分析
2022-11-30 06:48李健,毛可欣,王海荣,安淼,刘光柱,李国田
山西果树 2022年2期
关键词:猕猴桃
李健,毛可欣,王海荣,安淼,刘光柱,李国田

摘要:【目的】獼猴桃作为富含维生素的“水果之王”深受群众喜爱,但近年来多地发生冻害情况,给猕猴桃生产带来巨大损失,该研究针对猕猴桃冻害、生理指标、基因表达进行了分析,以便为猕猴桃抗寒性研究提供理论基础。【方法】对4年生‘泰山1号‘ 徐香等10个猕猴桃品种,于2月17日观察冻伤切面,于2021年3月15日和2021年3月25日,随机选取一个品种的3个植株,每个植株随机选取两个枝条,统计萌芽率。同时对生理指标丙二醛、可溶性糖、脯氨酸、电导率、超氧化物歧化酶(SOD)进行了测定。【结果】通过试验,从分子、表观层面分析了不同猕猴桃品种间抗寒性的差异,发现软枣、中华系猕猴桃抗寒性强于美味系猕猴桃,得到了适于山东泰山地区生产的‘泰山1号‘脐红‘金红50和‘龙成2号猕猴桃品种,部分美味系猕猴桃抗寒性太差,不适合山东地区种植。该研究可以通过生理指标、基因表达情况选择更为抗寒的猕猴桃品种,可以开发抗寒性分子标记,以快速筛选抗寒品种。
关键词:山东泰山;猕猴桃;抗寒性分析
文章编号:2096-8108(2022)02-0021-06中图分类号:S663.4文献标识码:A
Investigation and Analysis of Cold Resistance among Kiwifruit Cultivars in Taishan Area, Shandong Province
LI Jian1, MAO Kexin1,2, WANG Hairong1, AN Miao1, LIU Guangzhu3,LI Guotian1*
(1.Shandong Institute of Pomology, Taian 271000, China;
2.Shandong Agricultural University, Taian 271000,China;
3.Feicheng Peach Industry Development Center,Feicheng 271600,China)
Abstract:【Objective】 Kiwifruit, as the ‘king of fruits, which is rich in vitamins, is loved by the consumers. But in recent years, many reports have been reported about the freezing injury, which has brought great losses to kiwifruit yield. In order to provide theoretical basis for research on cold resistance of Kiwifruit, this study analyzed freezing injury, physiological indexes and gene expression of Kiwifruit.【Method】 Ten kiwifruit varieties, including four-year-old taishan No.1 and Xuxiang, were randomly selected on March 15, 2021 and March 25, 2021 to observe the frostbite section on February 17. Three plants of a variety were randomly selected on March 15, 2021 and two branches of each plant were randomly selected to count the germination rate. At the same time, the physiological indexes malondialdehyde, soluble sugar, proline, electrical conductivity and superoxide dismutase (SOD) were determined.【Results】The differences in cold resistance between different kiwifruit cultivars were analyzed from molecular and apparent levels. It was found that the cold resistance of Actinidia arguta and Actinidia chinensis was stronger than that of Actinidia deliciosa. The kiwifruit cultivars ‘Taishan 1, ‘Qihong, ‘Jinhong 50 and ‘Longcheng 2 suitable for growing in Taishan area of Shandong Province, part of the delicious department of kiwi cold resistance is too poor, not suitable for shandong planting.This study can select more cold-resistant kiwifruit varieties through physiological indicators and gene expression, and develop cold-resistant molecular markers to quickly screen cold-resistant varieties.
Keywords:Taishan area of Shandong; kiwifruit; cold resistance analysis
猕猴桃(Actinidia chinensis Planch.)富含维生素C,被誉为“水果之王”,属猕猴桃科(Actinidiaceae)猕猴桃属(Actinidia Lindl.)藤本植物。原产于中国,主要分布于N20°至N50°[1],我国猕猴桃自然资源丰富,种植广泛,深受群众喜爱。影响猕猴桃产量的因素主要有气候、病虫害、水肥条件等,其中冻害及早晚霜对猕猴桃生长带来严峻考验。此外,猕猴桃种间抗寒性参差不齐,同一品种在不同气候、栽培条件下其抗寒能力也存在差异[2],加之盲目引种、管理不当,南种北引往往造成严重损失。其中美味、中华猕猴桃两个主栽种主要分布在黄河以南,耐寒性较差,最低耐受-10 ℃左右[3]。随着近年来极端低温天气的增多,全国每年都有大批猕猴桃种植区遭受冻害,仅陕西省宝鸡市在1991-2016年间的冻害面积就达3.57万hm2[4]。
冻害发生在早春可以使幼叶、芽、花蕾发生坏死;冻害发生在深秋可以造成树干及新枝冻伤,降低植株休眠时对低温的耐受力;冻害发生于冬季能使枝干断裂、休眠芽冻伤,最终导致其无法正常萌发[5],冻害不仅对猕猴桃正常生长造成影响,还会造成减产,甚至大面积死亡。2021年1月上旬遭遇了近-20 ℃极端低温天气,本研究以山东省果树研究所泰东试验基地‘泰山1号‘徐香等北方主栽品种为材料,于2020年11月-2021年4月取材试验和调查,对猕猴桃冻害、生理指标、基因表达进行了分析,为猕猴桃抗寒性研究提供理论基础。
1材料与方法
1.1试验材料
供试植株为4年生‘泰山1号‘徐香等共10个猕猴桃品种,位于山东省果树研究所泰东科研基地,东经116°20′~117°59′、北纬35°38′~36°28′,年平均降水量697 mm,属温带半湿润大陆性季风气候,土壤为中性偏酸沙壤土,有機质质量分数1%,碱解氮质量分数19~43 mg/kg,磷质量分数21~34 mg/kg,钾质量分数32~68 mg/kg。转录组与生理指标测定为‘泰山1号‘徐香‘脐红‘龙成2号4个品种,直径为0.5~0.8 cm的1年生新枝作为样本,2020年11月15日至2021年4月15日共取6次。
1.2试验方法
极端低温出现于2021年1月6日-7日,冻伤切面观察于2月17日。萌芽率统计于2021年3月15日和2021年3月25日,随机选取一个品种的3个植株,每个植株随机选取两个枝条。萌芽率=萌发芽数/总数×100%。
生理指标测定:将取得的枝条液氮速冻后用磨样机磨成粉末,按0.1 g分装到不同离心管。丙二醛测定使用TCA法、可溶性糖测定使用蒽酮法、脯氨酸测定使用茚三酮法。电导率测定使用雷磁公司的DDS-307型电导率仪。超氧化物歧化酶(SOD)测定使用格锐思生物公司的WST法试剂盒,货号G0101F。
基因表达热图数据来源于美吉生物转录组测序结果,合同编号:MJ20201116080。
2结果与分析
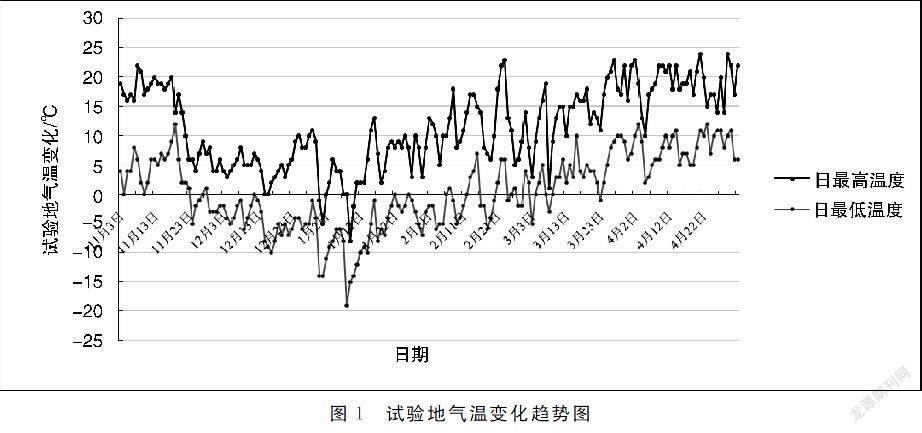
2.1试验地气温变化
由试验地气温趋势图(见图1)可以发现,11月下旬温度急剧下降,1月6日最低气温达到最低峰-19 ℃,1月7日最高气温达到最低峰-8 ℃,随后气温逐渐升高。采样时的日期温度分别为7~19 ℃、-10~2 ℃、-8~7 ℃、-2~8 ℃、4-16 ℃、7~19 ℃,其中1月15日所取样品为经历极端低温后的枝条。
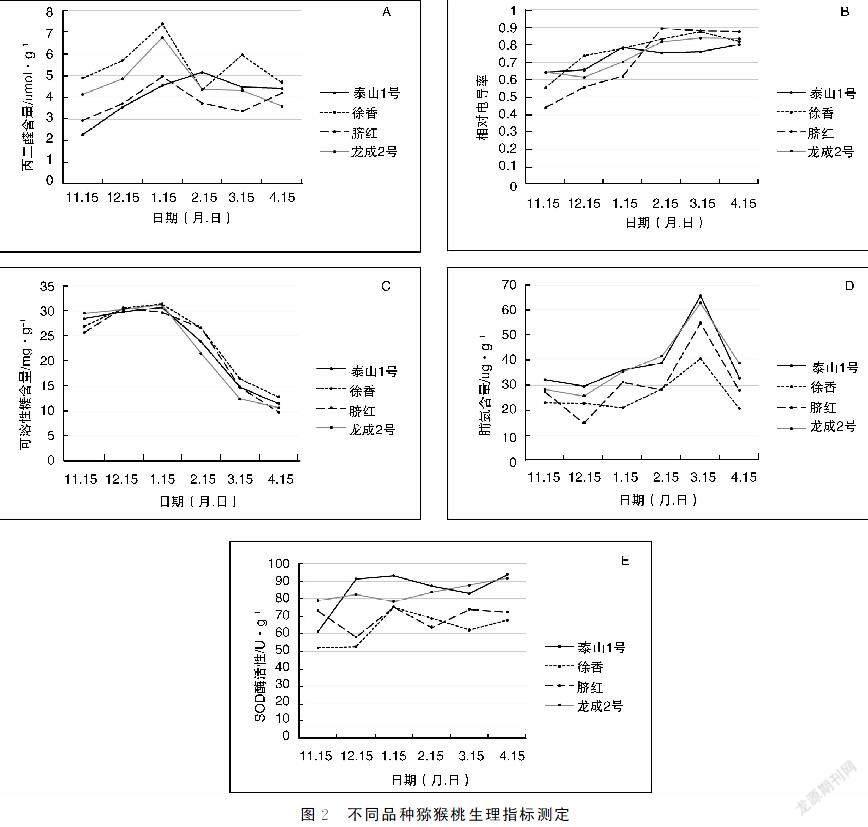
2.2猕猴桃不同品种生理指标测定
通过对6个时间点枝条生理指标丙二醛、相对电导率、可溶性糖、脯氨酸和SOD酶活的测定(见图2),丙二醛质量分数在1月15-2月15日到达顶峰随后下降,相对电导率总体呈上升趋势。可溶性糖含量缓慢上升至1月15日,随后一直下降,降至峰值的0.3~0.4,脯氨酸质量分数在3月15日达到顶峰,随后下降。SOD酶活在11月15日-1月15日和3月15日-4月15日两个时段内都有升高趋势。可以看出,在寒冷的气温下,猕猴桃膜损伤程度(丙二醛、相对电导率)上升,随着温度回暖膜损伤程度下降;另外,像可溶性糖、脯氨酸等抗冻性内溶物在低温下含量也有一定升高;过氧化物酶是植物体内重要的呼吸酶类,在低温胁迫后SOD酶活升高,提高抗逆性。由不同生理指标结果可以看出,‘泰山1号与‘龙成2号抗寒能力最强,其次为‘脐红,抗寒性最差的是‘徐香。
2.3猕猴桃不同品种抗寒相关基因表达分析
对试验基地内4个猕猴桃品种枝条进行转录组测序,选取了26个植物抗寒相关基因进行表达热图绘制(见图3),1-6列分别表示2020年11月-2021年4月。基因的表达模式在4个品种内基本一致,其中抗寒基因HT1-2、HT1-3在12月、1月低温胁迫下表达量提高,而转录因子CBF5、CBF6在降温过程中表达量有所下降,这与其低温下过多结合冷诱导基因COR有关。植物抗氧化系统中SOD2、SOD3、CAT、POD等基因在降温过程中表达量上升,温度回升后其表达量随之下降。乙烯应答元件组合蛋白AP2/ERF、参与ABA信号转导的PYL9在气温降低后表达量迅速提高,最高可达12.85倍。个别基因的表达模式在不同品种间有差异,调控维生素C质量分数的AsA基因在‘泰山1号‘徐香中低温下表达量升高,在‘脐红中变化较小,而在‘龙成2号中有降低的趋势;MYB44基因在低温下表达量提高,而在‘龙成2号中未检测到其表达,推测这与品种差异有关,‘龙成2号属于软枣猕猴桃,较中华、美味猕猴桃差异更大。
2.4极端低温下猕猴桃不同品种冻害调查分析由于试验地区1月6日左右遭遇了-19 ℃极寒温度,造成了猕猴桃不同程度冻害,于2月中旬进行了10个品种的冻害调查。用刀片切开树皮露出形成层进行观察后发现(见图4),‘徐香猕猴桃冻害最为严重,下部形成层发黑褐色,‘泰山1号和‘金桃受到轻微冻害,其余品种形成层呈现绿色。通过调查还发现,猕猴桃形成冻害部位主要是离地20 cm以内的下部,而地上中部则冻害较轻,顶部枝条受影响最小。
试验基地猕猴桃于2021年3月上旬开始萌动,3月15日和3月25日分别统计猕猴桃萌芽率,死亡率调查于4月30日(见表1)。由结果可以看出,3月25日较15日萌芽率不同,正常条件下萌芽率应升高,但‘农大金猕‘米良1号‘金桃等品种萌芽率下降幅度较大,这与植株受冻伤有关,导致上部枝条供水供养不足。‘徐香冻害最为严重,萌芽率只有21.67%,而死亡率却高达63.64%,‘金魁冻害情况次之,这两个品种都属于美味系猕猴桃。抗冻能力较好的品种为‘泰山1号‘脐红‘金红50和‘龙成2号,此为中华系和软枣猕猴桃,适于山东泰山地区种植。
3讨论
从研究结果不难看出,猕猴桃品种间抗寒性存在明显差异,总体来看软枣、中华系猕猴桃抗寒能力高于美味系猕猴桃,但中华猕猴桃‘农大金猕冻害比美味猕猴桃‘米良1号‘翠香严重。渗透调节物质可溶性糖质量分数、脯氨酸质量分数可以一定程度反映植物抗寒能力的强弱[6]。有研究表明,低淀粉高可溶性糖的软枣猕猴桃抗寒能力高于其他软枣猕猴桃[7]。本研究中,4个品种可溶性糖质量分数都在低温冻害时上升,随温度回升后又下降,品种间差异不明显,但受冻害时脯氨酸质量分数‘泰山1号>‘龙成2号>‘脐红>‘徐香,确实反映了品种间抗寒能力。当然,丙二醛、相对电导率、SOD也能一定程度反映膜损伤程度,低温下植物SOD活性应该是先上升后下降,其下降到一定程度O2-会降低[8-9]。
植物抗寒性受多基因共同调控,随着植物抗寒性研究的不断深入,抗寒基因被不断地挖掘[10]。转录因子CBF是植物抗寒体系重要节点基因,可调节下游COR、BAM[11]基因,过表达CBF可使植株SOD活性提高低温抗性增强,同时也能影响赤霉素、生长素、脱落酸调节枝条休眠及落叶[12]。本研究中,CBF基因在气温降低过程中表达量有下降趋势,与其结合并调控下游抗寒基因密不可分。猕猴桃抗氧化系统SOD2、SOD3、CAT、POD17、AsA等基因在降温过程中表达量升高,与此同时生理指标SOD酶活也升高,抗氧化系统有效降低植物O2-浓度,从而调节细胞膜透性、增强细胞膜稳定性,保护植物体正常代谢[13]。植物生长素响应因子ARF参与植物生长发育各过程,最近许多研究表明植物ARF在环境胁迫响应也有重要作用[14],其可参与磷酸盐胁迫[15]、盐和干旱胁迫[16]等,在本研究中发现,在冷胁迫下ARF7表达量下降,温度回升后表达量提高。扩展蛋白(expansin),是一类提高植物细胞壁韧性的细胞壁蛋白[17],广泛参与植物生长各阶段、多种逆境胁迫[18],本研究中expansin LA1在低温过程中表达量上升,提高了猕猴桃抗寒性。猕猴桃抗寒是一个由多基因共同调控的复杂过程,基因表达的研究有助于抗寒品种的分子筛选工作。
猕猴桃不同品种抗寒性确实存在差异,结果显示,10个品种中抗寒性最弱的为‘徐香‘金魁‘农大金猕,抗寒性最好的品种为‘泰山1号‘龙成2号‘金红50和‘脐红。相对而言,山东泰山地区软枣、中华系猕猴桃的抗寒性要优于美味系猕猴桃,这与2009年郑州地区[19]、2016年泰山南麓[3]猕猴桃冻害调查结果相一致。虽然‘泰山1号中华猕猴桃抗寒性较强,但是今年在山东猕猴桃主产区淄博[20]却损失惨重,淄博市有近2 000 hm2猕猴桃[21],主栽品种‘泰山1号冻害严重,有的果园几乎全部冻死。分析其原因主要有以下几点:2020年冬季的极端低温,2021年初遭遇严重倒春寒,果园区块不同引起冷气聚集,地势不同受风力影响较大以及栽培管理设施有差别等。降低猕猴桃冻害损失需要一定措施,一是应该选育和引进抗寒品种;二是要规范化管理,水肥充足,注意排水,入冬前及时剪枝;三是由于猕猴桃冻害主要发生于离地10~20 cm处,在冬季可以进行高培土、树干刷白、捆扎稻草或秸秆;四是可以立挡风墙,一定程度减少冷风;五是在设施大棚中,防倒春寒可喷洒雾化水汽,防止昼夜温差大可在白天及时通风、夜间焚烧秸秆。
4结论
‘泰山1号‘脐红‘金红50和‘龙成2号等猕猴桃品种适宜山东泰山地區种植,部分美味系猕猴桃抗寒性太差,不适合山东地区种植。可以通过生理指标、基因表达情况选择更为抗寒的猕猴桃品种,可以开发抗寒性分子标记,以快速筛选抗寒品种。本研究为山东地区猕猴桃抗寒品种选育、猕猴桃抗寒性分子研究提供了一定理论基础。
参考文献
[1]Lin M M, Fang J B, Qi X J. iTRAQ-based quantitative proteomic analysis reveals alterations in the metabolism of Actinidia arguta[J]. Scientific Reports, 2017,(7): 1-11.
[2]孙世航.猕猴桃抗寒性评价体系的建立与应用[D].中国农业科学院,2018.
[3]黄永红,史修柱,李桂云,等.2016年泰山南麓猕猴桃冻害调查与分析[J].落叶果树,2016(6):17-19.
[4]李广文,贺瑶,李红娟,等.陕西宝鸡产区猕猴桃冻害发生规律调查[J].西北园艺(果树),2018(2):48-51.
[5]龚宏伟.红阳猕猴桃的冻害成因与防治措施探析[J].陕西农业科学,2013,59(5):134-136.
[6]Chen K, Arora R. Understanding the cellular mechanism of recovery from freeze-thaw injury in spinach: possible role of aquaporins, heat shock proteins, dehydrin andantioxidant system [J]. Physiologia Plantarum, 2014(150):374-387.
[7]Sun S, Fang J, Lin M, et al. Freezingtolerance and expression of β-amylase gene in two Actinidia arguta cultivars with seasonal changes [J]. Plants, 2020, 9(4):515.
[8]郭子武.设施油桃低温胁迫的研究[M].山东农业大学,2003.
[9]朱丽丽.柑橘应答低温胁迫的生理生化变化[M].华中农业大学,2008.
[10]姚晓华.植物抗寒相关功能与调控基因研究进展[J].中国农学通报,2013,29(27):141-147.
[11]Peng T, Zhu X, Duan N, et al. Ptr BAM1, a β-amylase-coding gene of Poncirustrifoliata, is a CBF regulon member with function in cold tolerance by modulatingsoluble sugar levels [J]. Plant, cell & environment. 2014(37):2754-2767.
[12]Artlip T, Mcdermaid A, Ma Q, Wisniewski M. Differential gene expression innon-transgenic and transgenic “M.26” apple overexpressing a peach CBF geneduring the transition from eco-dormancy to bud break [J]. Horticulture Research,2019(6):86.
[13]Sou M B. Reactive oxygen species and oxidative burst roles instress, senescence and signal transduction in plants [J].Current Science,2005(89):1113-1121.
[14]陈磊,李晨曦,江涛,等.毛竹生长素响应因子基因家族的鉴定及其在竹笋发育中的表达[J].福建农林大学学报(自然科学版),2016,45(6):662-667.
[15]Shen C, Yue R, Yang Y, et al.OsARF16 is involved in cytokinin-mediated inhibition of phosphate transport and phosphate signaling in rice (Oryza sativa L.) [J].Plos One, 2014, 9 (11): e112906
[16]Xu YX, Mao J, Chen W, et al. Identification and expression profiling of the auxin response factors (ARFs) in the tea plant (Camellia sinensis (L.) O.Kuntze) under various abiotic stresses [J]. Plant Physiol Biochem, 2016(98): 46-56.
[17]Cosgrove D J.Growth of the plant cell wall [J]. Nature Reviews Molecular Cell Biology, 2005, 6(11): 850-861.
[18]Zhang H, Ding Y N, Zhi J K, et al. Over-expression of the poplar expansin gene PtoEXPA12 in tobacco plants enhanced cadmium accumulation [J]. International Journal of Biological Macromolecules,2018(116): 676-682.
[19]齊秀娟,方金豹,赵长竹.2009年郑州地区猕猴桃冻害调查与原因分析[J].果树学报,2011,28(1):55-60.
[20]单保爽,侯乐明,巩文琼,等.山东淄博市称猴桃产业现状及发展建议[J].中国园艺文摘,2013(8):65-66.
[21]陈凤龙,乔峰,黄爱玲,等.淄博猕猴桃栽培技术要点[J].农业科技通讯,2021(4):302-304.
猜你喜欢
快乐语文(2021年36期)2022-01-18
今日农业(2021年11期)2021-11-27
今日农业(2020年19期)2020-12-14
动漫星空(兴趣百科)(2020年12期)2020-12-12
作文小学中年级(2019年9期)2019-10-14
好孩子画报(2017年12期)2018-01-06
妈妈宝宝(2017年4期)2017-02-25
创新作文(小学版)(2016年23期)2016-12-01
新农业(2016年13期)2016-08-16
新农业(2016年13期)2016-08-16
