
鸡肉参蜜腺和传粉机制研究
2022-11-16 07:29周丽贞
蜜蜂杂志 2022年10期
周丽贞
(丽江师范高等专科学校,云南 丽江 674100)
植物传粉包括了自花传粉和异花传粉,其中异花传粉是有花植物最普遍的传粉方式[1]。对于植物传粉的研究包括了花粉和柱头生物学、虫媒授粉的演化、多样性的地理趋势、植物-授粉者的对应关系等[2],开展传粉生物学研究在探索自然、生物资源利用、农林业生产和生物多样性保护等实践上有重要意义[3]。鸡肉参(Incarvillea mairei(Levl.)Grierson)属于紫葳科(Bignoniaceae)角蒿属植物,为喜马拉雅-横断山脉的多年生草本植物[4],花冠较大,颜色为紫红色,根为肉质根,具有一定的观赏性与药用价值[5-6],研究其传粉机制对引种栽培、原生地资源保护利用等具有重要意义。在自然界中,花粉与柱头的结构演变出纷繁复杂的传粉适应性和传粉策略[7],在这些适应性中,有一些是通过柱头的运动和形态变化来适应恶劣环境和授粉昆虫稀少的限制,以提高异花授粉的几率[8]。鸡肉参具有敏感性二裂片柱头,这类柱头在物理性碰触后会闭合,不满足授粉要求则又能够再次打开,闭合速度因种而异;如果由于授粉而闭合柱头,多数不再打开[9-11]。一些研究指出这类柱头的运动可能是为了避免或延迟自花授粉[12-13]。研究通过对鸡肉参雌蕊及蜜腺的微观观察,以及花粉-胚珠数(P/O)比值,蕾期、半开期、平展期、凋谢期的柱头运动性,旨在探索鸡肉参潜在的传粉机制和二裂片柱头运动的意义。
1 材料与方法
1.1 实验材料
鸡肉参新鲜花朵和雌蕊不同部位样品取自丽江甘海子,海拔3020 m,离丽江师范高等专科学校实验室约31km。样品花粉-胚珠数比值实验、体式解剖镜观察、固定液配制、保存等均在师专实验室进行,后期处理和扫描电镜观察在中科院昆明植物所完成,柱头运动性实验在甘海子进行。
1.2 实验方法
1.2.1 解剖镜实验
取新鲜花朵装入保鲜袋,带回实验室用体式解剖镜解剖观察。
1.2.2 扫描电镜实验
1.2.2.1 样品固定
样品的固定采用了磷酸缓冲戊二醛固定液,戊二醛终浓度为2.5%。磷酸缓冲液分为甲液和乙液,甲液为0.2 mol/L磷酸氢二钠溶液,乙液为0.2 mol/L磷酸二氢钠溶液。采样当天取24.5 mL的甲液加入25.5 mL乙液混匀,即为0.2 mol/L的磷酸缓冲液(pH=6.8),并加入10 mL 25%戊二醛水溶液,用蒸馏水定容至100 mL混匀,分装入5 mL指型试管待用。在采样过程中使用了冰袋和泡沫箱,以减少温度的影响。
样品根据开花程度,分为蕾期、半开期、平展期、凋谢期四个时期分开采样,并且每个时期分别以柱头、子房、子房基部三部分取样。取样时,用双面刀片迅速切取样品放入事先标记的装有固定液的试管中,盖好盖子并放回泡沫箱中。待所有样品取好后,将样品带回师专实验室,以第一个样品采样开始计时至6 h,依次将所有样品更换新鲜配制的磷酸缓冲戊二醛固定液,放入4℃冰箱保存。后期样品也采用冰袋与泡沫箱运送。
1.2.2.2.样品处理和观察
样品采用梯度浓度的乙醇溶液脱水,其中初始处理浓度为30%,处理2 h后转入45%和60%乙醇溶液中,处理时间均为30 min,然后转入70%的乙醇溶液中处理过夜,再依次转入80%、90%乙醇溶液中,每阶梯停留时间为15 min,最后以无水乙醇处理3次,每次处理时间10 min。将脱水后的样品进行临界点干燥,上台并进行扫描电镜观察与拍照。
1.2.3 花粉及P/O值测定
根据孙鹏等[14]的方法,随机取10个花蕾,分别取其花药,用2 mL 0.1mol·L-1HCl溶液在60℃恒温水浴中处理1 h,吸5μL于载玻片上,滴1μL I2-IK溶液观察计数[15]。重复取样观察6次,其中染色深黑色和浅黄色的花粉粒分开记录。取子房在解剖镜下观察并记录胚珠数。
1.2.4 柱头运动行为观察
分别根据花开程度,对蕾期、半开期、平展期、凋谢期的花朵进行柱头运动性实验,对柱头裂片内侧进行物理性碰触,同时记录完全闭合时间以及闭合后再次打开的时间,并对观测数据进行Excel统计分析。蕾期需要用刀片从花冠1/2处横切,将上部花冠轻轻的垂直取下以便观察柱头。
2 结果与分析
2.1 解剖镜观察结果
通过解剖镜观察,鸡肉参柱头等二裂,淡黄色,内具绒毛,由花柱与子房相连。子房绿色,在子房基部具有环状凸起,子房和基部环状凸起上均发现有液体(图1A-B),表明鸡肉参具有环状蜜腺,且子房上也可能分布有蜜腺,并以花蜜吸引授粉者传粉,但通过观察发现泌蜜量并不多。

图1.解剖镜图片(4×)(A:子房基部的环状蜜腺;B:正在分泌花蜜的子房外壁)。
2.2 扫描电镜观察结果
2.2.1 柱头结构
扫描电镜视野下,鸡肉参柱头裂片外部并不光滑,外表皮细胞特化为具有纵向皱褶纹路的表皮毛,在130倍数下观察,如鳞片状覆盖在柱头裂片外侧,其间散布有特化的气孔和蘑菇状细胞。气孔与子房基部环状蜜腺的泌蜜孔相似,蘑菇状细胞与子房外部的相似(图2A)。
柱头裂片内外侧差别巨大,与外侧不同,内侧密布指状表皮毛(图2A)。表皮毛自裂片边缘一直延展至与花柱连接处,靠外的指状表皮毛较细小,向心的指状表皮毛较粗大,当花粉粒落在裂片内侧指状表皮毛上,花粉粒萌发,花粉管自表皮毛细胞进入花柱头(图2B)。
2.2.2 子房外部结构
子房表皮细胞排列紧密,无气孔状泌蜜孔分布,但散布有蘑菇状细胞,多数为饱满的近圆球体,部分皱缩瘪塌(图2C-D)。结合解剖镜观察结果,推测这一类细胞可能是一类分泌细胞,具有分泌花蜜的能力。

图2.柱头和子房超显微结构。(A:柱头内外部结构对比(120×);B:花粉管从柱头内侧指状细胞中进入(1.00K×);C:子房表皮的显微结构(120×);D:子房上饱满的蘑菇状细胞(1.00K×)。
2.2.3 蜜腺
通过扫描电镜对子房基部的环状蜜腺进一步观察,发现上面分布有气孔状泌蜜孔。根据蕾期、半开期、平展期、凋谢期四个时期采集的样品发现,蕾期泌蜜孔紧闭(图3A),花朵半开至平展期泌蜜孔打开(图3B-C),凋谢期泌蜜孔被一些物质填塞(图3D)而失去开合和分泌物质的能力。
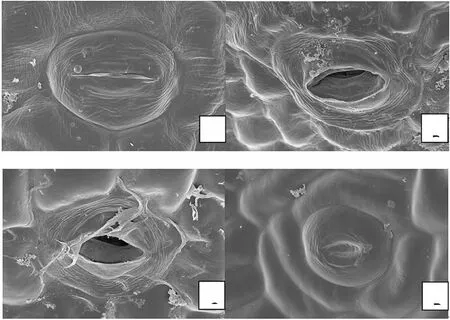
图3.不同开花时期的泌蜜孔变化(2.00K×)(A.蕾期;B.半开期;C.平展期;D.凋谢期)
2.3 花粉-胚珠比(P/O)及蕾期花粉活力
通过统计分析,鸡肉参单花总花粉粒为72300±4837.8064,胚珠数136.3684±5.6716,P/O约为530,根据分类,鸡肉参属于兼性异交型繁殖系统[16]。通过I2-KI染色,其中深黑色花粉粒为70340±5130.9224,占总花粉数的97.11%;浅黄色花粉粒1960±1043.6971,占总花分数的2.89%,说明在蕾期,大量花粉就已经成熟。
2.4 柱头的运动性
在实验过程中发现,蕾期柱头并未打开,在花冠取下后1h以上仍然闭合,凋谢期的柱头则保持闭合或打开而失去运动能力,仅半开期和盛开期具有运动能力,其实验结果如表1所示。

表1 柱头二裂片运动实验结果
在物理碰触下,半开期柱头的闭合时间比盛开期柱头闭合的时间短,且两者具有显著性差异P=0.0135(<0.05,df=9,10),表明柱头在半开期比盛开期对碰触更为敏感;而再次打开的时间,两者之间没有显著性差异P=0.1407(>0.05,df=9,10),表明在半开至盛开的时候,开花程度对柱头闭合后再次打开没有影响。
3 讨论
开花植物能够通过花内蜜腺分泌具有糖分的液体吸引并反馈给传粉昆虫,从而实现虫媒植物不同花朵之间的传粉,分泌花蜜的通道往往是蜜腺上特化的气孔,又称为泌蜜孔[17-18]。泌蜜孔一般分布于蜜腺上,在传粉昆虫采食花蜜的时候,虫体的绒毛接触花粉和柱头而得以传粉。通过体式解剖镜和扫描电镜的观察,鸡肉参蜜腺为子房基部的环状蜜腺,上面分布有泌蜜孔,且随着花朵的开放与凋谢,泌蜜孔表现为:闭合-开放-被物质填塞而失去分泌花蜜能力的变化。
除了子房基部的环状蜜腺,在二裂片柱头外侧也发现了类似泌蜜孔的结构,子房与柱头则还存在蘑菇状细胞,这些可能与泌蜜和吸引昆虫有关,如建兰花蜜的分泌是经由表皮毛分泌的[19]。花蜜是重要的传粉反馈物,泌蜜量和花蜜的甜度受到花的性别、移出量等影响[20-21],在一些植物的研究中则发现未被采食的花蜜会被再吸收利用[22-24]。植物分泌花蜜需要消耗大量的能量,它们往往需要在因花蜜产量增加而产生更多的受精种子和成本之间进行权衡[25]。结合体式显微镜观察,子房外壁有液体分泌,推测子房上的花蜜是经由蘑菇状凸起分泌而出,当凸起细胞的水分含量增加,细胞变得鼓胀饱满,细胞渗透压增加(图2D),在内部压力和传粉昆虫给予的外部压力(如攀爬、舔舐等)共同作用下,带有糖分的液体从凸起基部的缝隙中渗出以反馈给传粉者,当花蜜分泌到一定量时,凸起细胞皱缩(图2C)。如此,在没有授粉昆虫访问时,可以避免多余的花蜜分泌,减少能量的消耗。
一般认为,高海拔植物由于授粉昆虫的限制而更多的进行自花授粉,但对Eritrichium nanum的研究表明,自花授粉随着海拔的增加而降低[26]。根据P/O(≈530)表明鸡肉参为兼性异交型植物,且蕾期花粉已经基本成熟,但二裂片柱头没有打开,在半开期,柱头更为敏感,而随着盛开到凋谢,柱头运动能力降低直至失去运动能力,这可能是鸡肉参在高海拔的环境条件下为了最大限度保证异花授粉的原因。
4 结论
鸡肉参属于兼性异交型植物,为提高异花传粉的几率,表现出蕾期花粉粒多数成熟,而二裂片柱头闭合,半开期裂片触碰闭合速度比盛开期快等特点。花蜜是鸡肉参主要的传粉反馈物,雌蕊基部具有环状蜜腺,上面分布有气孔状泌蜜孔,从蕾期至凋谢期,泌蜜孔表现出“闭合-开放-被物质填塞而失去分泌花蜜能力”的变化。雌蕊柱头亦有类似泌蜜孔的结构分布,而柱头外侧和子房上则分布有蘑菇状特化细胞,并且子房具有分泌花蜜的能力。综合以上分析,鸡肉参由于受到高山环境传粉昆虫稀少的限制,为提高异花传粉几率,减少自身能量消耗,演化出了特有的花部结构和访花昆虫之间的协作机制。
感谢:在此对谷志佳老师和赵文正老师以及在实验过程中热忱帮助我的老师、朋友和同事表示感谢。
猜你喜欢
山东林业科技(2022年3期)2022-07-14
新疆农业科学(2022年9期)2022-02-21
西北农林科技大学学报(自然科学版)(2021年5期)2021-05-31
舰船科学技术(2021年1期)2021-03-09
广西植物(2020年13期)2020-09-12
山东工业技术(2019年12期)2019-05-30
草堂(2018年11期)2018-11-13
阅读与作文(小学低年级版)(2016年10期)2016-10-12
江苏农业科学(2016年2期)2016-04-11
