
盐度及种间相互作用对海三棱藨草、互花米草萌发及生长的影响
2022-11-11 13:26史欢欢于振林王天厚汪承焕
生态学报 2022年20期
王 倩,史欢欢,于振林,王天厚,汪承焕
华东师范大学生命科学学院,上海 200241
群落构建理论认为,区域物种库内所有物种的功能性状经过局域生物和非生物因子的筛选后形成该局域的物种集合[1]。在植物的不同生活史阶段,影响局域物种更新和空间分布的机制存在差异。群落形成的早期,种子散布和萌发决定了先锋物种在空间上的大致分布[2],而环境因子对于萌发生态位的筛选进一步推动了群落的发育[3—4]。更新阶段是植物生活史的关键环节,包括种子散布、萌发、幼苗建成等过程[5],探究更新过程在群落构建中所起的作用是我们理解物种多样性形成机制的关键。
长江口盐沼湿地具有明显的环境梯度,植物群落大部分处于演替早期,物种组成简单,是研究植物群落构建机制的理想系统[6]。盐度是长江口盐沼湿地最重要的环境因子之一[7]。盐生植物能够耐受一定程度的盐环境,成为维持当地生态系统的基础。然而,高浓度的盐分会干扰植物吸收水分,对胚和发育中的幼苗形成离子毒害,影响细胞正常代谢活动[8—9]。植物在不同生长发育阶段对盐胁迫的耐受能力不同,种子萌发期对于盐分最为敏感,直接关系到个体能否成活,进而影响种群的建立[10—11]。此外,种间相互作用是影响植物群落构建的另一个关键因素。由于人类活动的影响,生物入侵对各类生态系统的威胁不断加剧,入侵种与土著物种间的竞争作用正受到越来越多的关注[7,12]。
互花米草原产于北美东海岸,引入我国后迅速凭借其高繁殖力和耐盐性大肆扩张,成为滨海湿地生态系统的重要入侵物种。在长江口崇明东滩,互花米草的入侵挤占了中、低潮位海三棱藨草等先锋物种的生存空间,造成土著植物生态位的收缩[13—14]。目前,相关研究主要关注入侵种和土著种的种群分布、生理特性的比较,以及群落的空间结构及多样性等方面,对于更新过程在群落构建中所起的作用还不够了解。本研究聚焦于崇明东滩优势物种互花米草和海三棱藨草在生活史早期萌发阶段的相互作用及对盐度的响应,并与生活史中后期阶段的生长与竞争状况作对比,以较为全面地理解盐沼植物群落构建机制,为东滩湿地的保护工作提供一定的参考。具体的科学假设是:盐度、种间相互作用对互花米草与海三棱藨草不同生活史阶段的影响存在差异,因此盐沼湿地植物群落的构建机制比我们原先的理解更为复杂。
1 材料与方法
1.1 研究区域
崇明东滩是长江口规模最大、发育最为完善的河口型潮汐滩涂湿地,位于上海最东端(31°26′—31°37′N、121°46′—122°02′E)。崇明东滩的表层土壤盐度通常在2‰—15‰之间[6],咸潮入侵时则可达20‰[15]。20世纪90年代,互花米草被引进崇明东滩后快速扩散,使原有植物群落格局发生了巨大变化。目前,崇明东滩北部高潮带以芦苇和互花米草的单一群落以及混生群落为主,中潮带为互花米草群落-海三棱藨草混生群落,南部以海三棱藨草、藨草以及糙叶苔草等莎草科植物群落为主[16]。
1.2 目标物种
海三棱藨草是崇明东滩先锋群落的优势土著物种,互花米草则是当地重要的外来入侵物种。本实验所用种子均于实验上年秋季取自崇明东滩,其中海三棱藨草种子从东滩当地种子商户处购得,互花米草种子采集自北八滧互花米草-芦苇混生群落,所有种子均放置在4℃冰箱内浸水保存并定期换水。
1.3 实验设计
1.3.1萌发实验1
2015年4月,开始进行互花米草与海三棱藨草的种间竞争萌发实验。在直径9cm的培养皿底部覆盖双层滤纸,随机分为淡水、盐水两种处理,分别在培养皿中滴加6mL的纯水或1%NaCl溶液,使滤纸均匀浸湿。每种处理下设置互花米草单种、海三棱藨草单种、互花米草+海三棱藨草混种3组物种组合,每组各4个重复。单种组每皿放置30粒种子,混种组每皿每个物种各放置30粒种子。种子洗净消毒后放入培养皿,加盖置于光照培养箱中。
1.3.2萌发实验2
2017年3月,开始进行互花米草与海三棱藨草的种间竞争机制实验。分别取互花米草和海三棱藨草种子各50g置于2个500mL的锥形瓶中,加入400mL蒸馏水密封,在30℃恒温摇床中震荡48h,加入适量NaCl制成盐度为1%的互花米草、海三棱藨草种子浸泡液,同时配置1%NaCl溶液,置于4℃冰箱内保存。在直径9cm的培养皿底部覆盖双层滤纸,将互花米草种子分成3组:单种组、互花米草+海三棱藨草混种组、互花米草+海三棱藨草种子浸泡液组,其中前两组在培养皿中滴加6mL的1%NaCl溶液,浸泡液组则滴加等量海三棱藨草种子浸泡液;海三棱藨草种子同样设置单种、互花米草+海三棱藨草混种、添加互花米草种子浸泡液三组处理,每组处理各4个重复。单种组和浸泡液组每皿放置20粒种子,混种组每皿每个物种各放置20粒种子。种子清洗消毒后放入培养皿,加盖置于光照培养箱中。萌发结束后,将培养皿烘干,加入30mL纯水充分浸泡种子和滤纸后,使用便携式盐度计(SANXIN 5052 salt)测定溶液盐度。
两个萌发实验中,光照培养箱(一恒MGC- 100)的设置均为白天(08:00—20:00)温度30℃,光强度100%;晚上(20:00—次日08:00)温度18℃,光强度0%。每隔24h补充适量水分维持滤纸湿润,每天统计记录各物种种子的萌发数。
1.3.3盆栽实验
2019年4月,开始进行互花米草与海三棱藨草的盆栽生长竞争实验。准备1L花盆(底径12.5cm,高度17cm,口径16cm)填入适量河沙,每盆加入Osmocote缓释肥2g(15- 10- 12- 2MgO-TE),分别放入装有淡水或1%NaCl溶液的周转箱中。设置互花米草单种、海三棱藨草单种及两物种混种3组处理,每组处理4个重复。单种组每盆均匀放入种子30粒,混种组每个物种各放30粒,在花房室温下培养,每隔5d更换周转箱内溶液。前3周每天统计萌发的种子数,2019年10月收获全部植物,测定每个物种的密度、高度和地上生物量。
1.4 数据分析
萌发率GP(%)=萌发种子数/实验种子数×100%
萌发速度Gv=∑(Gi×Di)/∑Gi,式中Gi:第i天发芽数;Di:天数。萌发速度用平均萌发时间来衡量,时间越短,萌发速度越快[17]。
采用多因素方差分析检验盐度及种间作用对萌发实验中互花米草与海三棱藨草萌发率、萌发速度的影响,以及对盆栽实验中互花米草萌发率、高度、地上生物量的影响。采用单因素方差分析检验不同处理对互花米草与海三棱藨草萌发率、萌发速度及最终盐度的影响,后续检验使用Tukey′s HSD test。盆栽实验中海三棱藨草在1%盐度处理下没有萌发,故采用独立样本t检验分析淡水处理下种间作用对海三棱藨草萌发率、高度、地上生物量的影响。所有数据分析使用SPSS 23.0进行,α=0.05,制图软件采用SigmaPlot 10.0。
2 结果
2.1 盐度及种间作用对互花米草、海三棱藨草种子萌发的影响
盐度、种间作用及其交互效应对互花米草种子的萌发率均无显著影响,但1%盐度显著降低了互花米草种子的萌发速度,而种间作用及交互效应对萌发速度则无显著影响(表1,图1)。
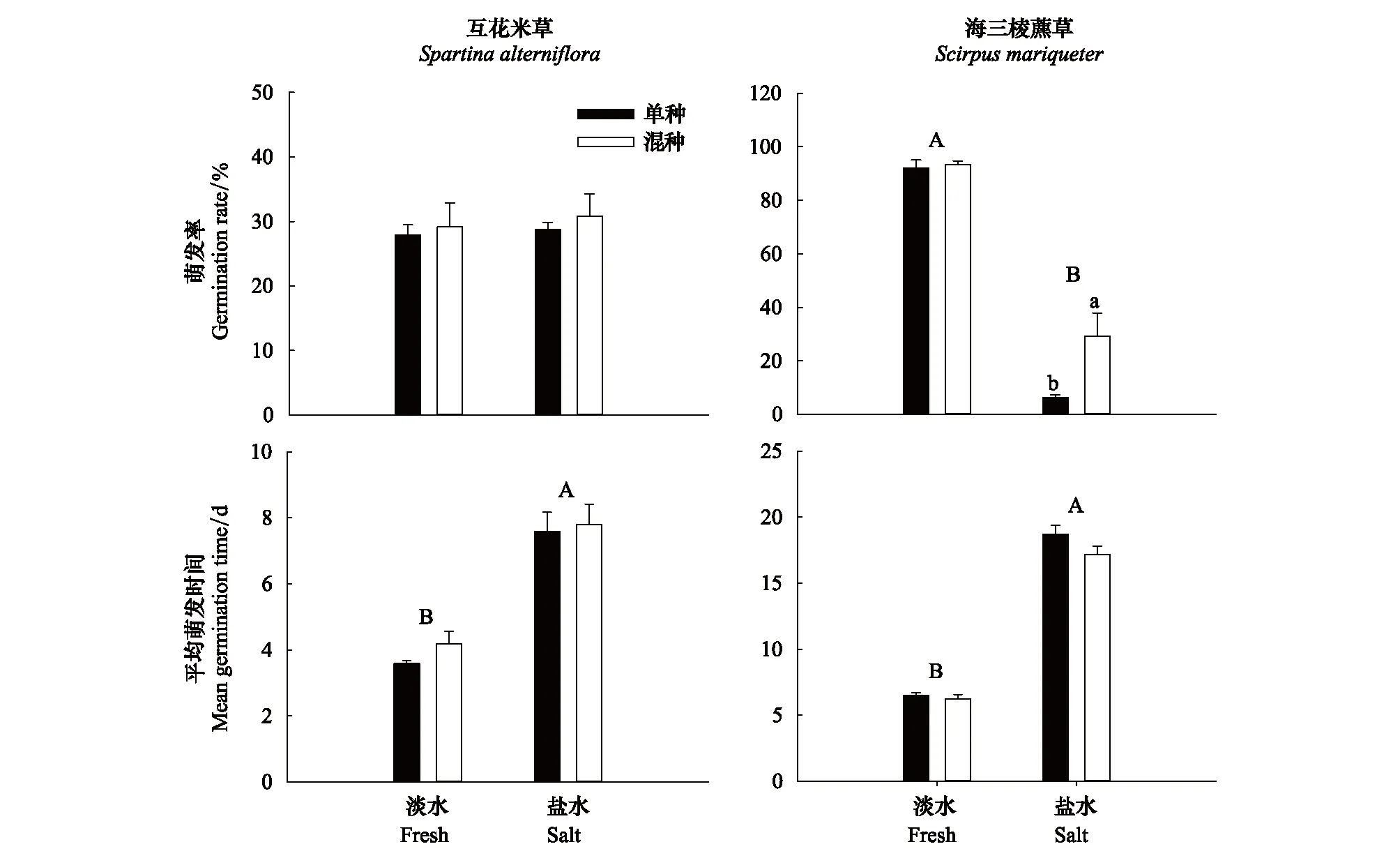
图1 互花米草、海三棱藨草种子在不同盐度及种间作用处理下的萌发情况Fig.1 Germination of Spartina alterniflora and Scirpus mariqueter seeds under different salinity and interspecific interaction treatments 不同大写字母表示不同盐度处理间存在显著差异,不同小写字母表示同一盐度处理下单种、混种组合间存在显著差异
盐度、种间作用及其交互效应对海三棱藨草种子的萌发率均有显著影响(表1)。1%盐度处理下海三棱藨草种子的萌发率显著低于淡水组;种间作用关系也受到盐度调控,淡水处理下互花米草种子对海三棱藨草的萌发几乎没有影响,而在1%盐度处理下,互花米草对海三棱藨草种子的萌发率有显著促进作用。海三棱藨草种子的萌发速度受到1%盐度处理的显著抑制,而种间作用及交互效应则无显著影响(表1,图1)。
2.2 竞争者种子及其浸泡液对互花米草、海三棱藨草萌发的影响
不同种间作用处理下,互花米草种子的萌发率、萌发速度均存在显著差异(表2)。混种处理组的互花米草种子萌发率显著高于单种组及添加海三棱藨草种子浸泡液处理组,后两者之间没有显著差异;单种组萌发速度最快,平均萌发天数显著低于浸泡液处理组,而混种组介于两者之间(图2)。
不同种间作用处理下,海三棱藨草种子的萌发率、萌发速度也存在显著差异(表2)。混种组的海三棱藨草种子萌发率显著高于单种组及添加互花米草种子浸泡液组,但萌发速度却显著低于另两组(图2)。其中浸泡液处理下的4个重复组中仅有1组萌发,无法比较各处理组间的差异,t检验结果表明单种组与混种组的萌发速度(平均萌发时间)存在显著差异(图2,P=0.011)。
互花米草、海三棱藨草种子萌发培养皿内溶液的最终盐度在不同处理组之间也存在显著差异(表2),混种组的最终盐度均显著低于单种组及添加竞争者种子浸泡液处理组(图2)。
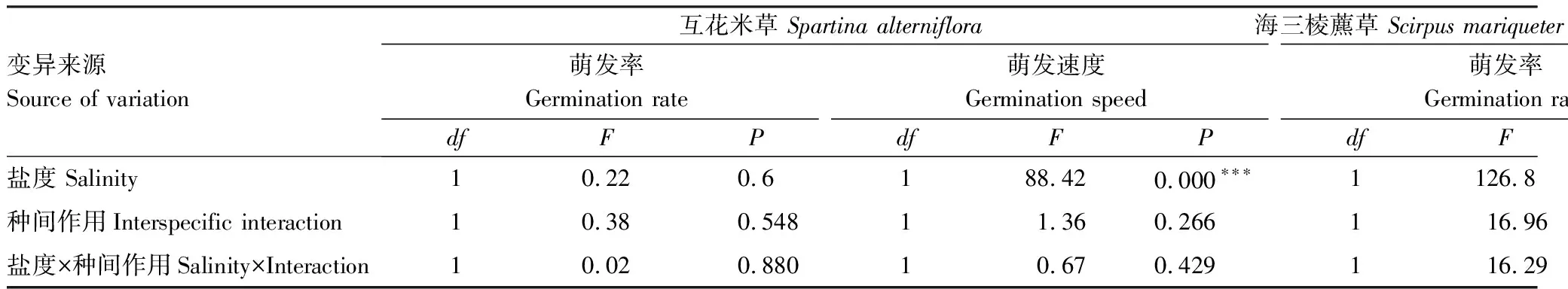
表1 盐度、种间作用对互花米草、海三棱藨草种子萌发影响的方差分析

表2 种间作用对互花米草、海三棱藨草种子萌发和培养皿溶液最终盐度影响的方差分析
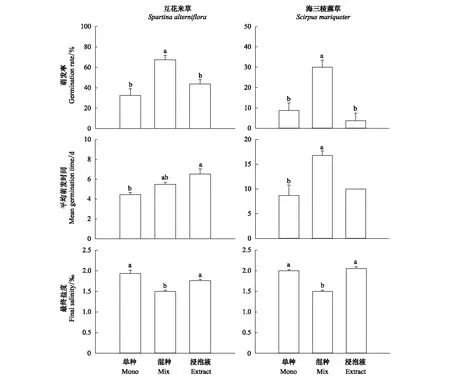
图2 不同种间作用处理下互花米草、海三棱藨草种子的萌发情况和培养皿溶液最终盐度Fig.2 Germination of Spartina alterniflora and Scirpus mariqueter seeds and final salinity in petri dishes under different interspecific interaction treatments 不同小写字母表示不同处理间存在显著差异
2.3 盆栽环境下互花米草、海三棱藨草的萌发及生长
与培养皿实验结果不同,盆栽环境下,盐度对互花米草种子的萌发率存在显著效应(表3)。1%盐度显著降低了互花米草种子的萌发率,而种间作用及交互效应则无显著影响(表3,图3)。盐度和种间作用对互花米草萌发速度的影响不显著,但二者的交互效应显著(表3),淡水处理下互花米草混种时的萌发速度高于单种,而1%盐度处理下则正好相反(图3)。
盆栽环境下,盐度对海三棱藨草种子萌发的抑制作用更为强烈,1%盐度处理下海三棱藨草完全没有萌发。在淡水处理下,种间作用降低了海三棱藨草的萌发率(图3),但效应不显著(t检验,P=0.247)。由于海三棱藨草种子萌发较晚且持续了数月,未能完整统计其萌发日期,故无法准确计算其平均萌发时间。
收获时,盐度、种间作用及其交互效应对互花米草的平均高度和地上生物量均无显著效应(表4)。混种组中互花米草平均高度和地上生物量都低于单种组,两者的差异在淡水中更大,但都不显著。海三棱藨草对盐度十分敏感,1%盐度处理下没有萌发。淡水处理下,混种组的平均高度、地上生物量也低于单种组,但差异不显著(图4)。

表3 盐度及种间作用对盆栽环境下互花米草种子萌发率和萌发速度影响的方差分析

图3 盆栽环境不同盐度和种间作用处理下互花米草、海三棱藨草种子的萌发情况Fig.3 Germination of Spartina alterniflora and Scirpus mariqueter seeds under different salinity and interspecific interaction treatments in pot experiment不同大写字母表示不同盐度处理间存在显著差异
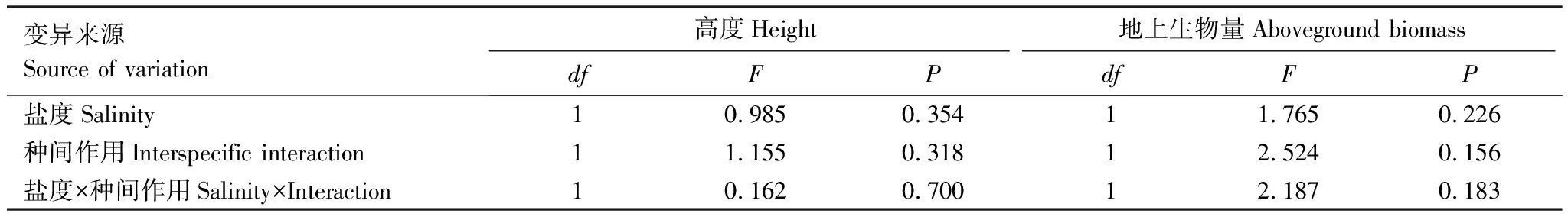
表4 盐度和种间作用对收获时互花米草生长表现影响的方差分析
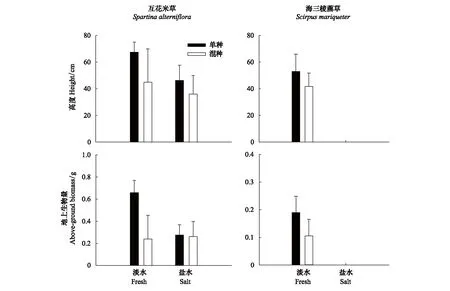
图4 盆栽环境不同盐度和种间作用处理下互花米草、海三棱藨草收获时的生长表现Fig.4 Growth performance of Spartina alterniflora and Scirpus mariqueter under different salinity and interspecific interaction treatments in pot experiment
3 讨论
3.1 盐度对互花米草、海三棱藨草萌发及生长的影响
长江口崇明东滩湿地具有明显的环境梯度,发生咸潮入侵时,盐度可达20‰左右[15],因此盐度是决定这一区域植物群落格局的重要因子。研究发现,互花米草具有较广的盐度生态幅(2‰—30‰)[18],成体植株甚至可以耐受高达60‰的盐环境。在高盐度(15‰—30‰)环境下,互花米草种子的萌发相比芦苇、碱蓬等土著物种更具优势[19],但盐度对其萌发及生长仍存在一定的抑制作用。盆栽生长竞争实验中,互花米草在1%盐度处理下萌发率显著降低(图3),平均高度和地上生物量也有所下降(图4)。海三棱藨草对盐度更为敏感,李伟等[20]研究表明,互花米草的耐盐阈值为43‰,高于海三棱藨草存活的耐盐阈值21‰。种间竞争萌发实验中,海三棱藨草在1%盐度处理下萌发率显著降低(图1),而盆栽生长竞争实验中,海三棱藨草在1%盐度处理下则完全无法萌发。另一方面,萌发速度对于植物快速占领生境十分重要[21],个体大小的不对称性有利于先萌发的物种在竞争中占据优势。种间竞争萌发实验中,尽管互花米草和海三棱藨草种子的萌发速度在1%盐度处理下都显著减慢(图1),但在相同盐度条件下,互花米草种子的萌发速度均显著高于海三棱藨草(淡水处理下平均萌发时间:互花米草(4.19±0.66) d,海三棱藨草(6.24±0.58) d,t检验,P<0.01;1%盐度处理下平均萌发时间:互花米草(7.80±1.08) d,海三棱藨草(17.19±1.07) d,t检验,P<0.01)。因此,即使土壤盐度未超过海三棱藨草的耐盐阈值,在同等盐度条件下互花米草种子的萌发速度相比于海三棱藨草也更占优势,进一步增加了其幼苗建成阶段的竞争能力。互花米草一旦成功定居后,其竞争优势更加明显,且能够通过强大的无性和有性繁殖来扩大种群,极易造成更大规模的入侵[22]。
需要注意的是,海水的组成比较复杂,对种子萌发的影响可能与NaCl溶液之间存在差别。张丽平等[23]的研究发现,重金属镉与盐度存在交互作用,能够加强NaCl对互花米草的胁迫。此外,对九段沙互花米草及海三棱藨草的研究发现,高盐度环境下,氮、磷营养盐尤其是氮的添加能显著促进互花米草地上部分的生长,而在低盐度环境下,氮添加则能促进海三棱藨草种子的萌发[24]。管博等[25]详细比较了NaCl、NaHCO3及海水溶液对盐地碱蓬种子萌发的影响,发现离子浓度相近时,NaCl溶液与海水处理下盐地碱蓬的萌发情况不存在显著差异。这一结果表明,海水对盐沼植物萌发的影响主要是由于NaCl的渗透胁迫和离子胁迫造成的,其他组分的影响较小。因此,NaCl溶液处理能够较好地代表海水的影响。植物的表现是不同环境因子共同作用的结果,除了海水盐度之外,其他环境因子也会产生重要影响。例如,水淹时间与盐度的交互作用对海三棱藨草幼苗的叶绿素及SOD含量有显著影响[26]。在今后的研究中,需要更多关注多种环境因子对植物的综合效应,以全面了解真实海水环境及其他环境因子对植物表现的影响。
3.2 种间作用对互花米草、海三棱藨草萌发及生长的影响
种间作用是影响植物种群建成的另一个重要因子。然而,除了少数研究之外,对于种子萌发阶段的种间相互作用及其内在机制了解仍十分有限[27]。种间竞争萌发实验中,胁迫较强的盐水环境中混种使海三棱藨草萌发率显著提高,互花米草的萌发率也略有上升(图1),此时种间具有一定的促进作用。而在淡水环境中,种间作用对互花米草、海三棱藨草种子的萌发基本没有影响,表明种间作用的类型和强度会受到盐度的调控。种间竞争机制实验中,互花米草、海三棱藨草种子在添加竞争者浸泡液处理组的萌发率、最终盐度均与单种组相近,而两个物种混种组的萌发率则显著高于单种组,且萌发结束后培养皿中溶液盐度显著降低(图2)。上述结果表明,互花米草与海三棱藨草在萌发阶段的相互促进并非是通过化感作用实现的,可能是由于种子萌发时对盐分的吸收减弱了盐胁迫的影响。有研究表明,盐胁迫会导致种子在萌发过程中胚细胞吸水困难,造成种子休眠或延迟萌发,盐度过高时,则会破坏细胞膜的完整性,造成种子死亡[28]。在种子萌发初期,盐度对于两个物种都是主要的限制因子,萌发时种子能够吸收一部分盐,使渗透胁迫得以缓解,提高了各自的萌发率,而萌发率的提高又进一步增加了对盐分的吸收,导致环境盐度显著降低。萌发阶段种间促进作用的具体机制还需要进一步深入研究。
需要注意的是,盆栽实验的结果与培养皿实验存在差异,混种并没有显著提高互花米草和海三棱藨草种子的萌发率。这可能是由于在培养皿中萌发时,种子之间相距较近,相对密度较大,盐度对于种子的影响更为直接,且培养皿容积较小,局部微环境较易受到种子生命活动的影响。在花盆中萌发时,种子相对密度较小,对局部微环境的影响相对较弱,因此互花米草与海三棱藨草之间的促进作用不明显。盆栽实验和培养皿实验确实能提供一些有价值的信息,但是将相关结果推广到野外时,需要更加谨慎[29]。
3.3 植物生存对策在不同生长发育阶段的变化
种间关系是影响群落构建的重要因素,但以往的研究大多聚焦于成体阶段外来入侵物种与土著物种之间的相互作用。长江口湿地的野外操纵实验及同质园受控实验均表明,入侵物种互花米草的种间竞争能力大于海三棱藨草[30—31]。两物种共存时,互花米草的种间竞争强度大于种内竞争,而海三棱藨草的种内竞争强度大于种间竞争[12]。将互花米草移栽至崇明东滩海三棱藨草群落后,随着其生长与扩散,海三棱藨草的多度、盖度、生物量、种子和球茎产量显著降低[32]。在长江口九段沙湿地,互花米草的扩散也导致了海三棱藨草群落的分布面积急剧减小[33]。因此,互花米草成体阶段强大的竞争力是其在长江口海三棱藨草群落成功入侵的重要机制之一。
植物在不同生长发育阶段受到干扰的强度不同,对于外界因子的响应机制同样存在差异[22,34],因此种间竞争的重要性和强度会随之发生变化[35]。已有研究发现,互花米草种子对盐水中海三棱藨草种子的萌发有显著促进作用,更新阶段植物种间相互作用的类型及强度与生活史后期阶段并不完全一致,仅考虑成体阶段在群落构建中的作用是有局限性的[27]。本研究结果表明,崇明东滩群落建成早期阶段,环境干扰较大,胁迫程度较高,竞争作用较弱,种间关系倾向于相互促进。种间竞争机制实验中,竞争处理下种子对盐分的吸收可以减缓盐胁迫,促进海三棱藨草及互花米草种子的萌发,有利于各自种群迅速占据新生境。一旦成功定居,随着植物的继续生长发育,生物量不断提高,干扰带来的影响逐渐降低,植株之间对于土壤养分、空间等资源的争夺加剧,竞争的重要性和强度也随之增加[22]。盆栽生长竞争实验持续时间更长,因而结果有所不同。混种组互花米草和海三棱藨草的株高、生物量相比于单种组均有所下降,这表明两物种间产生了一定的竞争作用,但是差异并不显著。这可能是由于在本次实验中使用的材料是由种子萌发产生的实生苗,而非营养繁殖形成的无性系分株。有研究表明,互花米草的无性繁殖体定居成功率、生长、繁殖等相关指标均远高于有性繁殖[36]。
在潮间带环境中,潮汐运动引起的滩涂淤积过程不断营造出新的生境,植物扩散到胁迫较大的新生境后,萌发阶段可能存在种间促进作用。然而,当群落发育到一定规模后,种群的进一步扩张及对资源的争夺会导致种间关系朝着竞争的方向演变,最终形成一个个单优势种的群落斑块。总之,物种之间促进或竞争的关系在环境因子及自身生长发育阶段的共同影响下不断波动,并处于一种动态平衡之中。
3.4 盐沼植物群落的构建机制
植物分布格局及各物种的共存关系一直是群落生态学的核心议题[37]。群落演替的早期阶段受种子影响较大[38],种子的扩散、萌发在较大程度上决定了种群在空间上的分布,但生境中有限的生态位和特定的环境条件又会约束种群的进一步发展[39]。本研究主要探讨了盐度及种间作用对崇明东滩优势物种互花米草、海三棱藨草的影响。在群落形成的早期阶段,种子散布及盐度等环境因子决定了空间内物种的基本组成,随着群落继续发育,种间相互作用开始取代其它环境因子成为新的主导因子,决定群落的结构和物种组成[40—41]。 互花米草与海三棱藨草存在着一定的竞争关系,并形成了明显的竞争位序。在崇明东滩中低潮带光滩区域,萌发阶段海三棱藨草与互花米草种子之间的竞争效应较弱,能否成功在新生境定殖主要取决于种子到达该区域的能力及对盐胁迫和潮水冲刷的耐受能力。相比于存续时间不超过一年的互花米草种子库,海三棱藨草2—3年的种子仍具有萌发能力,增大了种群存活几率。然而,植物群落地上部分密集的植株对海三棱藨草种子具有较强的截留能力,使大量种子在中高潮带沉积,大大降低了光滩区域的种子供应[42]。此外,在成熟群落中,互花米草与海三棱藨草均主要通过营养繁殖进行更新,由于竞争的不对称性,个体更大的互花米草占据显著优势,并逐渐取代海三棱藨草成为优势物种。因此,入侵种互花米草对海三棱藨草的威胁主要来自于成体植株。
在对盐沼湿地进行修复的过程中,除了加大对入侵植物互花米草的治理力度外,还需关注土著植物的恢复。要提高恢复效率,应根据不同物种在萌发及生长阶段对于环境因子和竞争的响应特征,采取适当干预措施,增大土著植物种子萌发和幼苗定植的成功率。在中低潮带,可以撒播预萌发的海三棱藨草种子或移栽幼苗,同时密切监测互花米草种子扩散形成的新生斑块,尽早加以清除,防止其发展为成熟群落。在中高潮带,清除互花米草后,可移栽海三棱藨草、芦苇幼苗,并充分利用湿地土壤种子库的潜能促进土著群落的自然恢复[43]。在高潮带,除了清除互花米草的无性分株外,还应注意控制其种子的生产与散布,芦苇、碱蓬、碱菀等土著植物则以自然恢复为主。总之,在长江口盐沼湿地治理过程中,应密切监测防控互花米草的再度入侵,因地制宜促进海三棱藨草等土著植被的恢复,提高湿地生态保护效率[44]。
致谢:约热尼沙对实验给予帮助,上海崇明东滩国家级自然保护区对调查采样提供帮助,特此致谢。
猜你喜欢
河北渔业(2022年10期)2022-10-15
当代水产(2022年8期)2022-09-20
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
昆明医科大学学报(2022年2期)2022-03-29
文萃报·周二版(2022年10期)2022-03-19
考试与评价·高二版(2020年4期)2020-09-10
知识就是力量(2020年2期)2020-05-19
学校教育研究(2020年7期)2020-04-09
发明与创新·大科技(2019年2期)2019-06-26
美与时代·美术学刊(2017年3期)2017-05-22
