
干旱胁迫对药用植物马比木叶片光合特性的影响
2022-11-01 03:31高渐飞
信阳师范学院学报(自然科学版) 2022年4期
高渐飞,周 玮,孙 燕,李 浪
(1. 贵州科学院 贵州省山地资源研究所, 贵州 贵阳 550001; 2. 贵州工业职业技术学院 工商管理系, 贵州 贵阳 550008)
0 引言
光合作用是绿色植物制造有机物的重要途径,植物生长发育过程中可将光能转化为可存储的化学能,为植物的生命活动提供最初的营养[1-2]。水分是影响植物的生长发育状态及各种生理活动的重要因素之一,也是限制植物在自然界中分布和影响其生产力的一个重要因素。然而植物本身对水分的需要与其所在环境的水分条件经常处于矛盾之中,水分不足或过多都会影响植物的光合作用,限制其正常生长[3]。植物在受干旱胁迫影响后,其叶片光合作用会产生明显响应[4]。其主要表现有光合酶的活性降低,净光合速率显著降低,延缓植物的正常生长[5-6];气孔阻力增大和CO2交换速率下降,抑制了叶片的扩展及干物质的积累[7];胞间CO2浓度的变化,气孔导度和蒸腾速率逐渐降低,影响植物的生长发育[8]。因此,了解干旱胁迫下植物光合作用特性响应规律,对构建适合其生长的环境具有重要意义。
马比木(Nothapodytespittosporoides(Oliv.) Sleum.)为茶茱萸科假柴龙树属植物,又名海桐假柴龙树、公黄珠子、追风伞等。分布在贵州、湖南、湖北等海拔230~1 100 m的山谷、阴坡等太阳直射少的阴湿地带,其对环境要求相对严格,生长缓慢。分布地多为喀斯特石山区,土壤以黑色石灰土为主,生境较为脆弱,一旦破坏,难以恢复。马比木以根入药,民间常用于祛风除湿、理气散寒,主治风寒湿痹、浮肿疝气、关节疼痛等病症[9-10]。近年来从根中提取的喜树碱类药物在临床上广泛用于治疗原发性肝癌、胃癌等多种恶性肿瘤及艾滋病[11-13],使其成为极具价值的抗肿瘤药物资源;加之其喜树碱含量远高于喜树果(之前喜树碱主要从中提取),从而致其遭受严重无序采挖,种质资源毁灭性减少,现今已十分稀少,濒临灭绝[13-16]。当前,关于马比木研究主要在繁育技术[17-18]、喜树碱的提取及含量测定方法[14-15]、内生真菌研究[19-20]和药效成分分析[21]等几个方面。自然环境中马比木多分布在阴湿地带,其是否对土壤水分和光照强度有特殊要求,有待进一步研究。研究采用LI-6400光合仪测定马比木叶片在不同干旱条件、不同光照强度胁迫下的光合特性,分析其净光合速率、蒸腾速率、气孔导度等响应变化,探寻其适合的土壤水分条件以及光照强度,对进一步了解其对干旱适应性及开展规模化栽植具有指导意义。
1 材料与方法
1.1 实验材料与处理
供试马比木树龄7~8 a。野生植株原生长于贵州省万山区,2014年11月移植(苗龄2~3 a,地径0.3~0.5 cm,苗高25~40 cm)至贵州省贵阳市贵州科学院(贵州省贵阳市云岩区)楼顶露天栽植,容器为口径25 cm、高30 cm塑料钵,土壤为林下去除掉落物层的腐殖土。栽植后适时浇水,至2016年3月未见明显生长。随即移栽至覆盖85%遮阳网塑料大棚,至7月新稍超过20 cm,生长旺盛。随后定期修剪,树高控制1.4 m以下,2018年12月从塑料大棚取苗后移栽于32 cm×30 cm(口径×高)的树脂塑料花钵中,用林下腐殖土和黄壤(w/w=1∶1)栽培,置于贵州科学院实验室阳台,定期浇水,长势良好。2019年10月8日(苗龄7~8 a,地径3~5 cm)分批次喷洒充分浇足水,之后让其自然干旱;第二日起,每2日测定叶片的光响应曲线,同时记录土壤水分含量(体积含水量),选择其中6个水分梯度的光响应曲线进行分析。
1.2 测定项目及方法
土壤含水量:采用ML3-KIT便携式土壤水分速测仪(感应区域高度5.5 cm×直径7.0 cm)分别测定距主干3~5 cm的周边东南西北4个方向土壤含水量,取平均值。
光响应曲线的测定:设置红蓝光源模拟设定光合有效辐射(PAR)次为1 400、1 200、1 000、900、800、700、600、400、200、100、50、0 μmol/(m2·s),每株(共3株)选取长势和光照接近部位3片成熟叶片,利用LI-6400便携式光合系统测定各处理的光响应曲线。测量时间为9:30—12:00,采用开放式气路,大气CO2浓度保持在394~402 μmol/mol。机器自动记录净光合速率(Pn)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和气孔导度(Gs)等参数,设置读数时间3 s,每次记录3~5个数值。
1.3 数据处理与分析
采用Origin 2018绘制不同土壤水分条件下各光合指标数据(Pn、Ci、Tr和Gs)和水分利用率(water use efficiency,WUE;Pn/Tr)的变化图,进一步分析其内在规律。采用Excel 2016对弱光条件下光合速率进行直线回归,得到光补偿点(light compensation point,LCP)和表观量子效率(the apparent quantum efficiency,AQE),根据Pn-PAR光响应曲线以及弱光时回归直线与最大净光合速率直线交点估计光饱和点(light saturation point,LSP)[22-23],并对所需数值进行显著性分析。
2 结果与分析
2.1 干旱胁迫对净光合速率的影响
在不同干旱程度下,马比木叶片净光合速率在PAR为0~200 μmol/(m2·s)之间呈快速上升趋势,200~600 μmol/(m2·s)辐射段上升趋势变慢,之后逐渐达到极大值4.09~7.16 μmol/(m2·s),见图1。在干旱过程中,净光合速率Pn先下降后逐步升高再迅速降低,当土壤含水量至10.5%时达最大值7.16 μmol/(m2·s);随之,当含水量减小为9.6%时,Pn值迅速下降至最小值4.09 μmol/(m2·s)。土壤水分31.3%~10.5%之间,随着干旱进行Pn逐渐升高至最活跃状态,继续干旱胁迫后Pn值迅速降至最低;土壤含水量为10.5%时,马比木可能进入适度的水分亏缺范围,可视其为耐干旱的极值。

图1 土壤水分对马比木净光合速率的影响Fig. 1 Effect of soil moisture on net photosynthetic rate of N. pittosporoides
2.2 干旱胁迫对胞间CO2浓度的影响
干旱胁迫对胞间CO2浓度Ci的影响见图2。
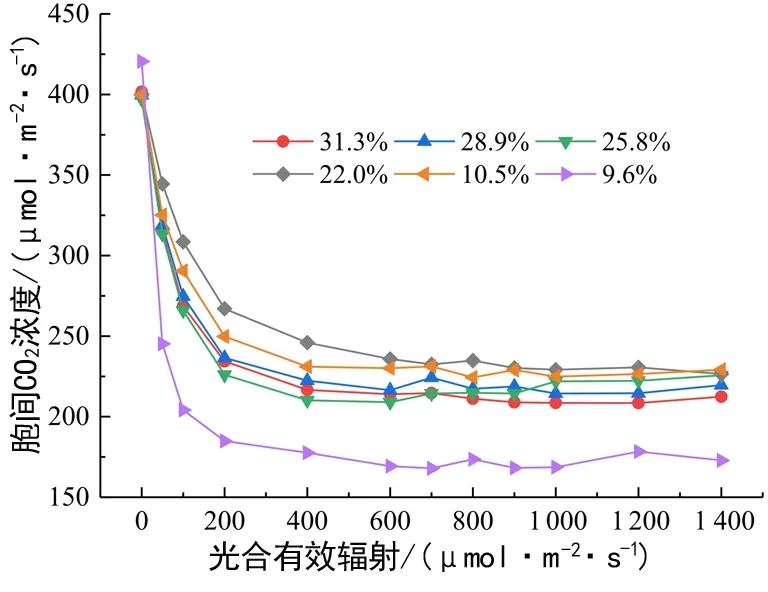
图2 土壤水分对马比木胞间CO2浓度的影响Fig. 2 Effect of soil moisture on the internal CO2 concentration of N. pittosporoides
PAR 在0~200 μmol/(m2·s)范围内呼吸作用所积存的CO2浓度消耗极快,200~600 μmol/(m2·s)之间Ci下降趋势变缓,PAR >600 μmol/(m2·s)后,Ci变化趋于平稳。土壤含水量在31.3%~22.0%之间,随着干旱进行Ci总体上呈现上升趋势至最高;之后出现回落,至9.6%时Ci陡降至最低(约175 μmol/(m2·s))。土壤含水量10.5%及以上的5个实验组之间的Ci均值无显著性差异(P>0.05),而9.6%实验组与其他各组的Ci均呈现差异显著(P<0.05);表明对于Ci值来说,22.0%是其光合活跃点,而10.5%是耐干旱极值点。
2.3 干旱胁迫对蒸腾速率的影响
图3中所示,随着干旱胁迫的进行,蒸腾速率Tr整体变化呈下降趋势,但随着辐射强度的增加呈现缓慢上升趋势。土壤含水量28.9%实验组Tr变化最快,9.6%实验组Tr下降最慢,31.3%实验组Tr偏弱。进一步看,PAR在0~100 μmol/(m2·s)范围时,除了9.6%实验组,其余各组的Tr均减小;PAR在100~600 μmol/(m2·s)之间,各组的Tr均呈逐步增加趋势;在600~900 μmol/(m2·s)辐射段,除9.6%实验组,其余各组Tr均呈现先上升后下降的趋势,这可能由气孔开合度引起,表明在此辐射范围内气孔变化较活跃。此外,该区间变化趋势与叶面温度变化基本一致,说明蒸腾速率与叶面温度相关,其随温度升高而增大。

图3 土壤水分对马比木蒸腾速率的影响Fig. 3 Effect of soil moisture on the transpiration rate of N. pittosporoides
2.4 干旱胁迫对气孔导度的影响
从图4可以看出,随着干旱进行,气孔导度Gs整体呈现出先升高后降低的变化趋势。
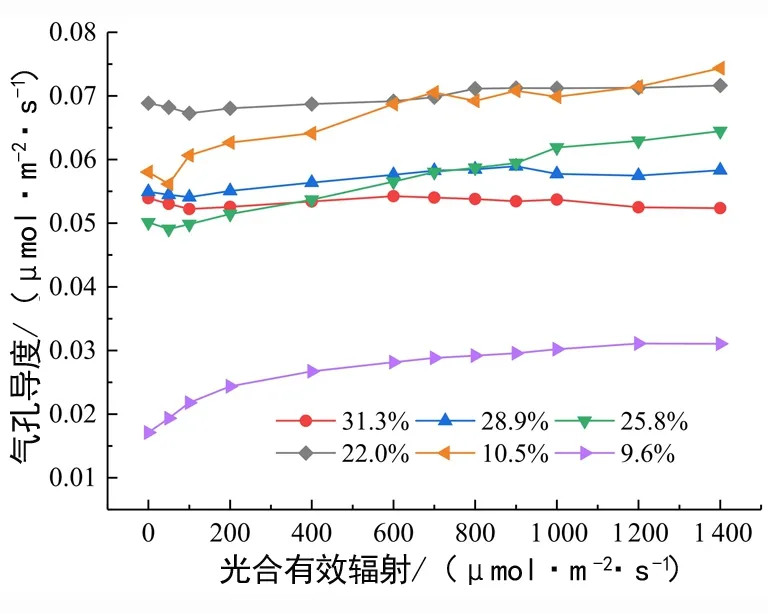
图4 土壤水分对气孔导度的影响Fig. 4 Effect of soil moisture on the stomatal conductance of N. pittosporoides
当PAR强于100 μmol/(m2·s)时,Gs开始随着PAR的增加而升高,而后呈平稳趋势。PAR <600 μmol/(m2·s)时,土壤含水量22.0%时Gs最大;PAR 在600~1 400 μmol/(m2·s)之间,含水量10.5%实验组的Gs已升至接近最高,至PAR为 1 200 μmol/(m2·s)时其超过22.0%实验组至最高值。而土壤含水量9.6%实验组Gs一直明显低于其他组,其中与10.5%实验组相比更是表现出显著性差异(P<0.05),说明10.5%是保持光合活性的耐旱极值,10.5%~22.0%是气孔导度活性较佳土壤水分。
2.5 干旱胁迫对水分利用效率的影响
图5为水分利用率(WUE)变化趋势图。

图5 土壤水分对水分利用效率的影响Fig. 5 Effect of soil moisture on water use efficiency of N. pittosporoides
由图5可知,低含水量WUE值较高,土壤含水量为10.5%时WUE值最大(约11 mmol/mol),土壤含水量为28.9%时,WUE值最小(约6 mmol/mol),两组之间的WUE值之间存在显著性差异(P<0.05)。在PAR为0~200 μmol/(m2·s)范围内,WUE快速升高,随后变化相对平缓。与前几个光合指标变化趋势相比,土壤含水量25.8%实验组WUE高于22.0%实验组。整体上看,高土壤含水量的2个实验组(28.9%和31.3%)WUE<较高的2组(22.0%和25.8%)WUE<较低实验组(10.5%);而9.6%实验组WUE几乎与31.3%实验组持平,虽然其光合效率低,但水分利用效率相对高。土壤含水量10.5%是6组实验中水分利用最高效的,表明适当干旱可以提高马比木的水分利用效率。
2.6 不同干旱胁迫的光响应参数
表1中,在干旱胁迫过程中,最大净光合速率(取平稳变化阶段的均值)呈现“减-增-减”的趋势,在土壤含水量10.5%时达到最大值6.91 μmol/(m2·s);22.0%时次之,为6.69 μmol/(m2·s);含水量低至9.6%时,最大净光合速率出现最小值3.97 μmol/(m2·s)。说明马比木的光合作用有其适合的土壤水分范围,且对干旱响应较为迅速。
随着干旱的进行,表观量子效率(AQE)整体呈减小趋势,值得注意的是,土壤含水量至10.5%时AQE增大至0.039,后再减小至0.028,依然是9.6%实验组与其余实验组间有显著性差异(P<0.05)。光补偿点(LCP) 先减小后增大,整体呈现“V”的变化趋势,其中土壤含水量9.6%实验组LCP值最大,达3.85 μmol/(m2·s);25.8%实验组LCP接近于0。光饱和点(LSP)呈“减-增-减”的趋势,最大值出现在22.0%实验组为187.61 μmol/(m2·s),其次是10.5%实验组为173.45 μmol/(m2·s)。整体上,干旱胁迫对马比木光补偿点与光饱和点变化影响较明显,土壤含水量25.8%时对弱光利用能力更强,10.5%~22.0%对强光利用能力更强。
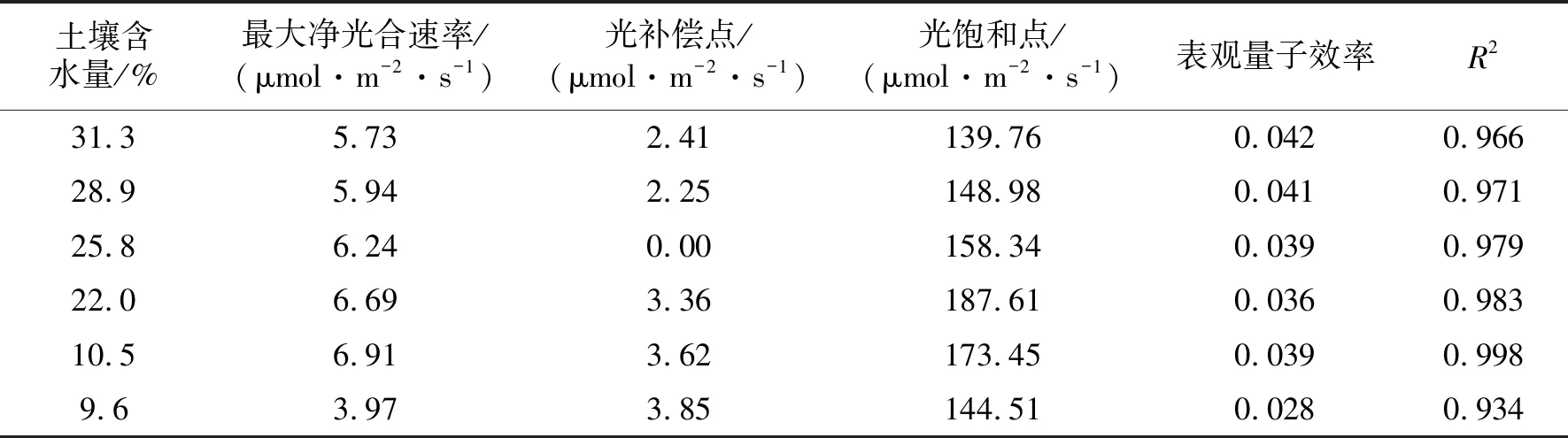
表1 不同土壤水分的马比木光响应参数Tab. 1 The value of light response parameter in different soil moisture of N. pittosporoides
3 讨论
通常情况下,水分会直接影响植物的生长,水分不足或过多都会影响到植物的光合作用[3],如何平衡植物生长与水分含量之间的关系将是植物适宜生长环境研究的重要环节。在马比木干旱胁迫中,实验设置6个不同土壤含水量来探讨马比木在干旱胁迫下光合指标的响应。
从各指标响应图看,Pn、Ci、WUE等3个光合指标主要呈现3个辐射阶段的响应,分别为弱光阶段(小于200 μmol/(m2·s))、中强光阶段(200~600 μmol/(m2·s))和强光阶段(600~1 400 μmol/(m2·s))。弱光阶段,各实验组Pn与WUE增加最快,相反,Ci则快速降低,其结果与苏金等[24]对紫珠的光合特性的研究结果一致;且随着干旱进行,指标增加(降低)幅度增大,此结果也在陈志成等[23]对珍珠油杏的研究中体现;说明弱光条件下马比木对干旱的应激响应更为强烈,一定程度上可以缓解干旱对植物造成的不利影响[25]。中强光阶段,6个实验组的Pn、Ci和WUE变化缓慢,是弱光阶段与强光阶段的过渡期,指标变化趋势基本与强光辐射阶段一致。强光阶段,3个指标基本平稳,且随干旱进行,呈现先增后减趋势。土壤含水量10.5%实验组Pn大于其余各组,22.0%实验组次之,而9.6%实验组的Pn最低,且与其余各组有显著性差异(P<0.05);Ci值变化中,10.5%和22.0%实验组降低缓慢,稳定时保持较高值;在WUE变化中,10.5%实验组对水分的利用率最大,22.0%~25.8%实验组次之,28.9%~31.3%与9.6%实验组处于低值;表明马比木在适度干旱条件下水分利用率提高[26],且土壤含水量在10.5%~25.8%之间能维持较高WUE。综合3个光合指标对干旱响应趋势,土壤含水量10.5%~22.0%是马比木光合生理活动最活跃区间[9,27],而10.5%的土壤水分是马比木的耐旱极值,22.0%是其光合活跃点。
在实验有效辐射范围内,Tr与Gs均呈现逐渐上升的趋势,且随着干旱进行,Tr减小,而Gs却在土壤含水量为22.0%和10.5%时、强光下保持最大。Gs与Tr息息相关,一般Gs降低,Tr随之降低[26]。研究显示:干旱胁迫会影响气孔的开闭[28],适当干旱反而刺激气孔打开,光合作用增强,实现自我调节[16],但随着干旱加剧,马比木已不能通过气孔调节实现自我保护,因此在含水9.6%时气孔导度显著(P<0.05)区别于其余各组。经进一步分析,高含水量(28.9%和31.3%)和最低含水量(9.6%)的3实验组叶面温度较高(23.5 ℃~25.0 ℃),其次为25.8%实验组, 10.5%和22.0%实验组叶面温度最低,而气孔导度与此相反,表明叶面高温可能是导致气孔关闭的另一个重要因素。
从影响马比木光合作用的因素分析,一般研究中有两种因素:气孔因素和非气孔因素,而Ci和Gs是判断两种因素的主要依据[28-29]。通常来说,Ci降低且Gs同时减小时,可判断Pn变化是由气孔因素引起,反之,则光合作用主要限制因素为非气孔因素[29-30]。实验中,马比木的Ci与Gs变化恰好一致,说明气孔因素是影响马比木光合作用的主导因素。
光补偿点(LCP)是植物光合速率与呼吸速率相等时的光照强度,反映的是植物对弱光的适应能力,其值越低,对弱光的利用能力越强[31]。在马比木水分利用率的研究中,随着土壤含水率的降低,LCP先降低后增加,土壤含水量在25.8%时接近于0,表明此时对弱光的利用能力最强。光饱和点(LSP)是植物Pn不再增加时的光照强度,反映的是植物对强光的适应能力,LSP越大,证明植物对强光的利用能力越强[32]。研究中,高含水量(25.8%~31.3%) LSP值在150 μmol/(m2·s)左右,中等水平含水量(22.0%~25.8%) LSP值在162 μmol/(m2·s)左右,较低含水量(10.5%~22.0%) LSP值平均为188 μmol/(m2·s),而低含水量(9.6%)仅为149 μmol/(m2·s)。整个变化规律呈现先增后减趋势,说明随着土壤含水率降低,马比木对强光的利用率增加,但是低于10.5%后,对强光利用率受到限制。由上可知,土壤含水率会影响马比木对弱光和强光的利用能力,总体上马比木对强光的利用能力更强。表观量子效率(AQE)反映的是弱光条件下植物吸收和转换光能的能力。研究中,马比木的AQE值在0.028~0.044范围,且随着含水率的降低,AQE值逐渐降低,说明弱光条件下,高含水量有利于马比木对光的利用[33]。
4 结论
马比木在干旱胁迫下有一定程度的自我保护调节机制,能适应较干旱的土壤环境。在适度干旱条件下,其光合速率增大,水分利用效率亦增大。实验表明,土壤水分含量10.5%可能是马比木耐旱极值, 22.0%~25.8%之间较适合其生长。马比木对弱光的利用能力较强,对强光也表现出较好的适应性,光照生态幅度较宽,且在强光下亦没有出现光抑制现象。但强光长时间照射马比木叶片是否会出现光抑制现象还有待进一步实验,以及所引起的温度上升可能会对其光合作用产生较大影响,并可能是其多分布在阴坡的主要原因之一,也有待进一步研究。
猜你喜欢
农业工程学报(2022年5期)2022-06-22
长江丛刊(2020年13期)2020-11-19
婚姻与家庭·婚姻情感版(2019年6期)2019-06-03
数字通信世界(2019年10期)2019-02-14
江苏农业学报(2018年6期)2019-01-04
青少年科技博览(中学版)(2018年6期)2018-08-16
现代园艺(2017年23期)2018-01-18
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
自然资源遥感(2014年2期)2014-02-27
