
外源乙烯对盐碱胁迫下花楸胚胎萌发的影响
2022-10-31 07:19王雨童赵彩鸿王晓东林木遗传育种国家重点实验室东北林业大学林学院黑龙江哈尔滨150040
种子 2022年9期
王雨童, 赵彩鸿, 王晓东, 杨 玲(林木遗传育种国家重点实验室,东北林业大学林学院, 黑龙江 哈尔滨 150040)
花楸(Sorbuspohuashanensis)是蔷薇科苹果亚科花楸属落叶乔木,是集观赏、药用和食用价值为一体的高效益珍贵树种[1]。野生花楸主要分布于中国的东北、华北等地,朝鲜北部和俄罗斯西伯利亚东部也有少量分布[2]。由于花楸的园林观赏价值逐渐被人们重视,过度的滥采乱挖对中国东北东部林区花楸的野生资源造成了极大的破坏,部分地区的种群濒临消失,因此,采取有效的保护措施保存花楸种质资源并加快花楸的开发利用研究具有重要意义。经研究发现,花楸树种子具有深度或绝对的休眠状态,属于严格休眠种子[3]。在花楸种子播种前期对其进行一定的层积处理能够有效地提高种子发芽率[4-7]。用花楸种子的水浸液对白菜种子进行培养后发现,白菜种子萌发受到了明显的抑制,说明引起花楸种子休眠的主要原因是花楸的果肉、果汁、种皮和种胚中均含有明显抑制种子萌发的物质脱落酸(ABA)[8]。ABA是导致花楸种子休眠的另一重要因素[9]。因此,可以将花楸种子休眠归类为生理休眠。最新研究发现,外源乙烯能提高花楸胚胎的萌发率,在胚胎破裂过程中乙烯生物合成的增加使NO代谢途径中H2O2含量增加,从而调控胚胎的休眠和萌发[10]。
近年来,我国多数地区土地盐碱化越发严重,限制了作物的生长和发育[11]。研究表明,土壤中的盐碱成分作为影响植物生长和产量最常见的胁迫因素之一,从种子萌发到开花结实整个生长发育过程中造成植物发芽率低、发芽延迟、幼苗死亡率高、叶绿素含量减少和光合性能降低等影响[12]。盐碱胁迫会导致种子吸水不足,子叶中储藏的蛋白水解酶活性受到抑制,使呼吸作用受到抑制,种子难以萌发[13]。盐胁迫降低了种子发芽率,从而损害了许多物种幼苗的建立[14]。有研究发现,外源乙烯可以有效地改善盐胁迫对种子萌发的抑制作用[15],但对其调控机制尚不明确。于建等[16]研究发现,在植物种子萌发的过程中,乙烯作为植物体重要的激素和信号物质,也参与到植物体众多生理生化及生长发育过程,包括参与植物响应生物和非生物胁迫。盐碱胁迫对花楸胚胎萌发的影响尚不清楚,乙烯是否可以提高盐碱胁迫下花楸胚胎萌发率尚待研究确定。
本试验用乙烯处理花楸胚胎,并对外源乙烯影响盐碱胁迫下花楸胚胎萌发的作用进行观察,对胚胎的萌发率、平均发芽速度、发芽势、发芽指数等数据进行记录与处理。研究结果为明确外源乙烯促进花楸胚胎休眠解除的作用机制,阐明乙烯缓解盐碱胁迫下花楸胚胎的作用机制提供理论依据,并为花楸在盐碱地区推广栽培提供参考依据,为人工促进花楸种子萌发、有效保护和扩展花楸自然资源提供技术和参考。
1 材料与方法
1.1 萌发试验
1.1.1试验材料
2020年10月上旬自东北林业大学帽儿山实验林场成年花楸母树采集成熟浆果后用水选法调制种子[17]。将处于安全含水量(9%~10%)的饱满、纯净成熟种子放入塑封袋,于4 ℃ 贮藏于冰箱中备用。
1.1.2萌发试验方法
1) 种子前处理
试验前测定种子千粒重、含水量和生活力,然后在蒸馏水中(20±5)℃下吸胀48 h。将吸胀后种子用0.2%(v/v)NaClO溶液搅拌浸泡15 min,用无菌水冲洗至少3次,在冰上剥除种皮(花楸为无胚乳种子)挤出裸胚用于试验,裸胚获得方法同杨玲等[18]的研究方法。
2) 萌发试验
乙烯(ETH)处理试验。将胚胎放在3 mL (ETH浓度为138 μmol/L)浸湿的滤纸上吸胀,对照为直接在蒸馏水浸湿的滤纸上吸胀。所有试验胚胎均在直径9 cm的培养皿中(每皿30个胚)25 ℃,光照强度为60 μmol/(m2·s)(每天16 h光照)下萌发。对照和处理均设3个以上重复。
碳酸钠(Na2CO3)、碳酸氢钠(NaHCO3)处理试验。将胚胎分别放在3 mL(Na2CO3浓度为8.92 mmol/L )、(NaHCO3浓度为14.15 mmol/L )浸湿的滤纸上吸胀,对照和萌发条件均同上。
Na2CO3、NaHCO3分别与乙烯的组合处理试验。将胚胎放在3 mL Na2CO3与ETH混合液、NaHCO3与ETH混合液浸湿的滤纸上吸胀,对照和萌发条件均同上。
1.2 数据处理与统计分析
采用Excel 2003软件进行数据整理,采用Sigmaplot 12.5软件进行绘图,SPSS 19.0软件进行数据处理系统方差分析、Duncan多重比较和相关性分析。方差分析在p=0.05水平上进行显著性检验。平均数间统计学差异用邓肯多重比较法在p=0.05显著性水平上检验。各指标计算公式如下:
萌发率(%)=(胚胎萌发数/供试胚胎总数)×100%;
平均发芽速度(d)=∑(D×n)/∑n
式中:D为从种子置床起算的天数,有的规定置床之日为0,有的规定为1。n为相应各天的发芽数。
发芽指数=∑Gt/Dt
式中:Gt为发芽试验终期内每日发芽数,Dt为发芽日数,∑为总和。
发芽势(%)=(日发芽数/供试胚胎总数)×100%。
2 结果与分析
2.1 乙烯对盐碱胁迫下花楸胚胎萌发的影响
不同处理对花楸胚胎萌发的影响如图1所示。用Na2CO3、NaHCO3处理后的花楸胚胎的萌发率显著低于对照,外源添加乙烯处理后萌发率虽没有达到对照水平,但也显著提高。由图2可知,在萌发过程中(1~8 d)最先萌发的是乙烯处理,第3天即可萌发,最晚萌发的是NaHCO3处理(第6天),第8天时各处理的萌发率由高至低依次为ETH(60%)、H2O(43%)、Na2CO3与ETH组合(28.89%)、NaHCO3与ETH组合(18.89%)、Na2CO3(13.33%)、NaHCO3(6.67%),且各处理间差异显著(p<0.05)。

注:不同小写字母表示不同处理间差异显著(p<0.05)。下同。图1 各处理对花楸胚胎萌发率的影响Fig.1 Effect of each process on the germination percentage of Sorbus pohuashanensis embryo
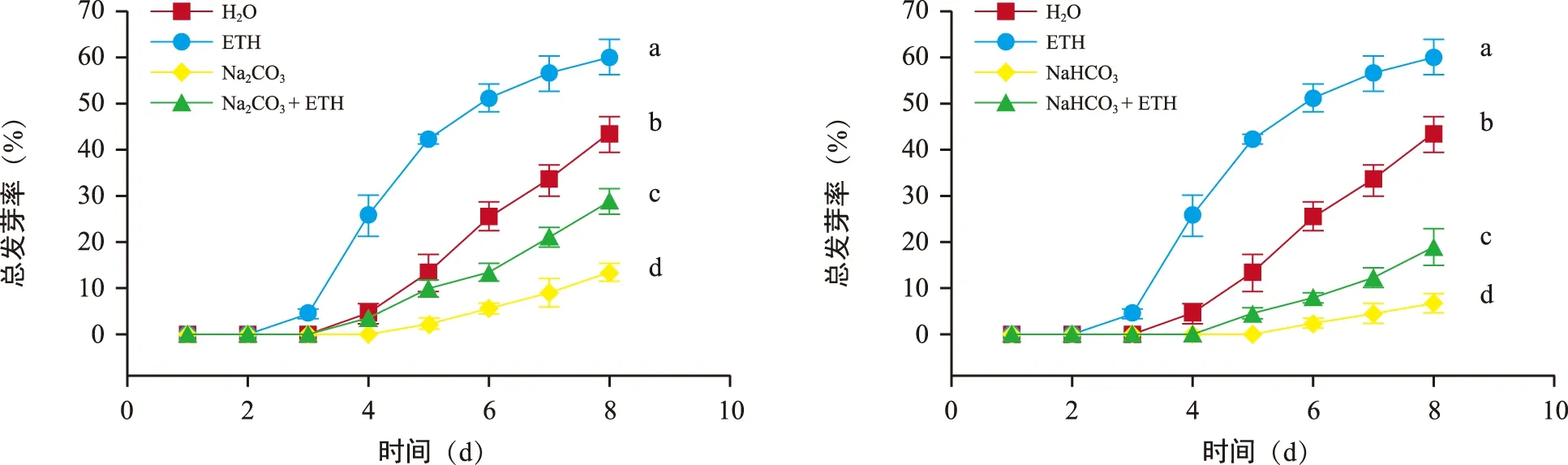
图2 各处理下花楸胚胎1~8 d的萌发率Fig.2 Germination percentage of 1 to 8 days under various treatments of Sorbus pohuashanensis
2.2 ETH对盐碱胁迫下花楸胚胎平均发芽速度的影响
平均发芽速度是指供试种子全部发芽所需要的时间,此指标可以反映种子的活力。由图3可知,ETH处理后花楸胚胎的平均发芽速度最低(4.98 d),显著低于其他处理。与对照相比,经Na2CO3、NaHCO3处理后花楸胚胎的平均发芽速度显著提高,Na2CO3为6.77 d、NaHCO3为7.17 d,说明胚胎萌发受到抑制。向Na2CO3、NaHCO3中添加外源ETH后,平均发芽速度降低,达到对照(6.25 d)水平,说明Na2CO3、NaHCO3对胚胎萌发的抑制作用减弱,胚胎萌发能力增强。
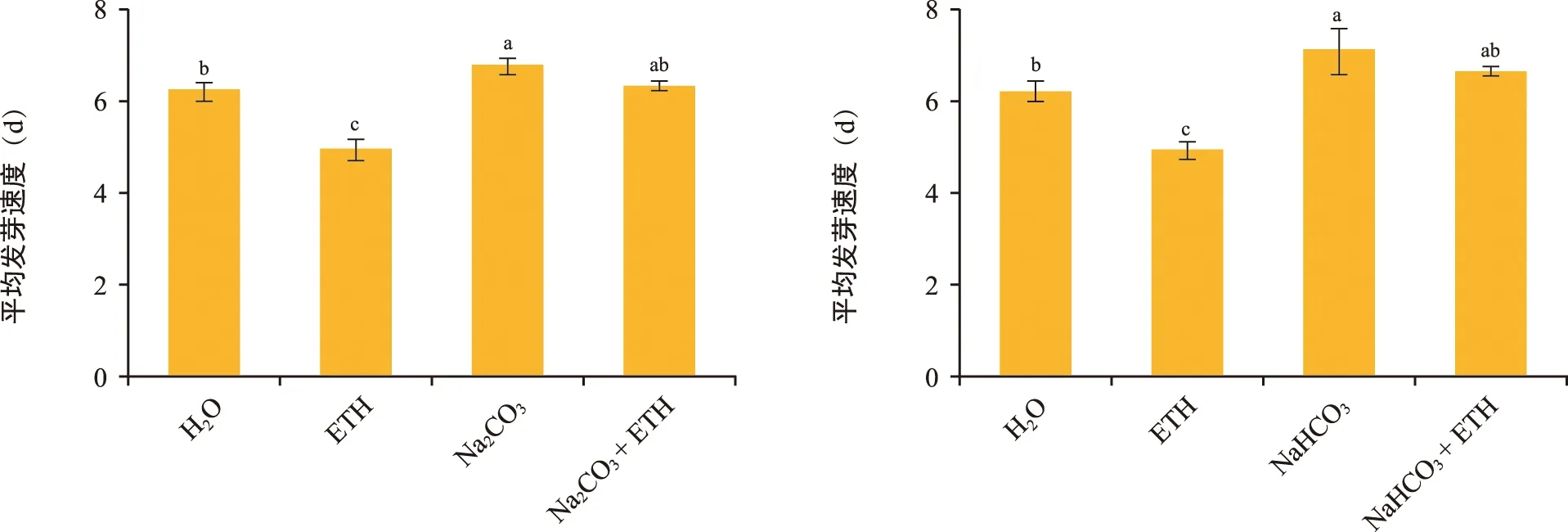
图3 各处理对花楸胚胎平均发芽速度的影响Fig.3 Effects of each process on the mean germination speed of Sorbus pohuashanensis embryo
2.3 ETH对盐碱胁迫下花楸胚胎发芽指数的影响
各处理中,ETH处理后的花楸胚胎发芽指数最高(图4),且各处理的发芽指数间均存在显著差异(p<0.05)。发芽指数最高的处理(ETH)是最低的处理(NaHCO3)的13倍多。由此可见,ETH不仅可以促进正常情况下的花楸胚胎萌发,还可以缓解盐碱胁迫对花楸胚胎的抑制作用。各处理下花楸胚胎1~8 d的发芽指数见图5。胚胎发芽指数在萌发过程中的变化是先增后降最后趋于稳定,说明胚胎在发芽中期活力最高。
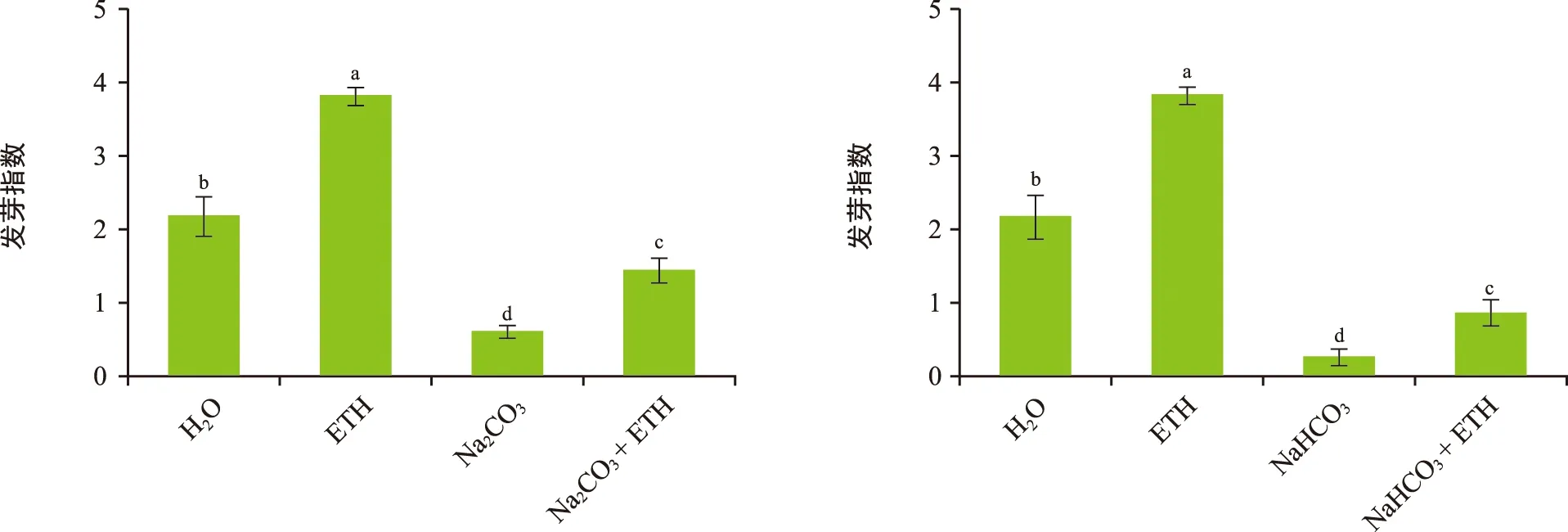
图4 各处理对花楸胚胎发芽指数的影响Fig.4 Effects of each process on the germination index of Sorbus pohuashanensis embryo
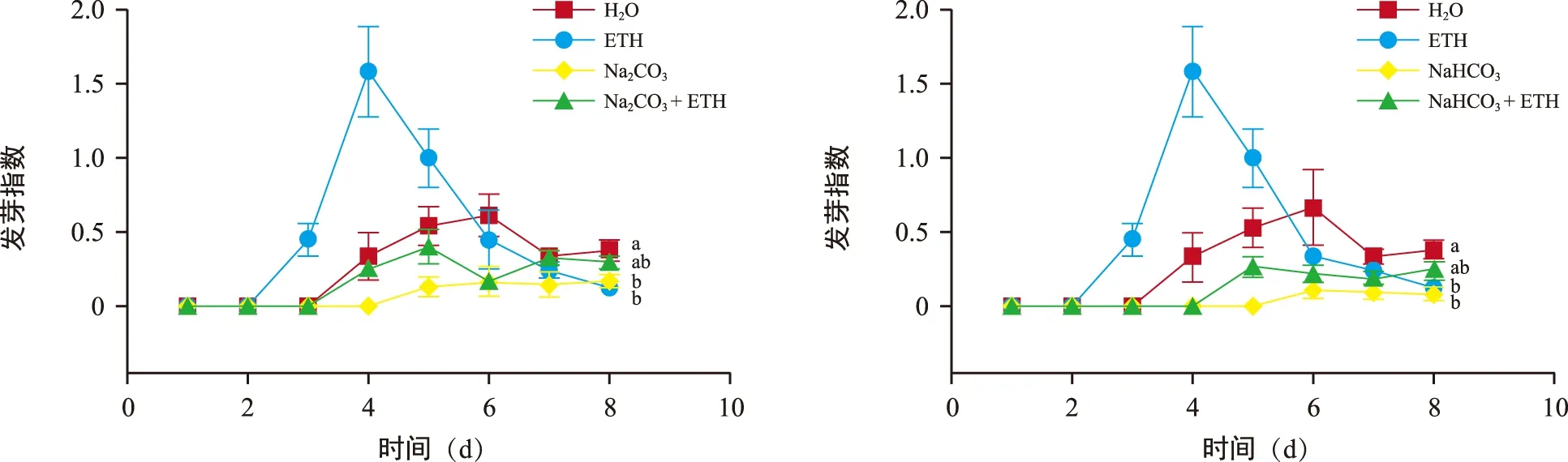
注:图中不同小写字母表示不同处理间差异显著(p<0.05)。图5 各处理下花楸胚胎1~10 d的发芽指数Fig.5 Germination index of 1 to 8 days under various processes of Sorbus pohuashanensis embryo
2.4 ETH对盐碱胁迫下花楸胚胎发芽势的影响
ETH处理对盐碱胁迫下花楸胚胎发芽势的影响见图6。ETH处理后胚胎的发芽势为24.44%,显著高于对照,Na2CO3、NaHCO3处理后胚胎发芽势显著低于对照。说明ETH可以促进花楸胚胎萌发,Na2CO3、NaHCO3抑制了花楸胚胎萌发。向Na2CO3、NaHCO3中加入ETH后花楸胚胎的发芽势分别提高40%和100%,但不显著,说明加入外源ETH可以缓解盐碱胁迫对花楸胚胎萌发的部分抑制作用。

图6 各处理对花楸胚胎发芽势的影响Fig.6 Effects of various processes on the germination potential of Sorbus pohuashanensis embryo
3 结论与讨论
3.1 Na2CO3、NaHCO3处理抑制花楸胚胎萌发
Na2CO3、NaHCO3是东北地区盐碱土中的主要成分,具有较强碱性,且pH值很高。盐胁迫会抑制盐生植物[19]和甜土植物[20]种子的萌发,碱胁迫除了直接抑制种子萌发外,还可以通过光合作用对植物生长进行影响[21]。本研究发现,与对照相比,用Na2CO3、NaHCO3处理后花楸胚胎的萌发率分别降低了70%和85%,发芽指数分别降低了72%和87%,发芽势分别降低了62%和77%,平均发芽速度分别提高了8%和14%,均与对照差异显著,说明盐碱胁迫条件下花楸胚胎萌发会受到抑制。
3.2 外源ETH处理促进花楸胚胎休眠解除
ETH是农业生产中常见的植物生长调节剂,在植物生长发育过程中发挥重要作用。近年来,ETH具有打破种子休眠的作用,已经逐渐得到证实,如苹果(MaluspumilaMill.)[22]。此外,赵秋荣和杨湘虹[23]通过研究ETH的生物合成、代谢以及信号通路发现,ETH会参与到种子的休眠与萌发。本研究结果发现,ETH处理后胚胎的萌发率、发芽指数、发芽势等均高于对照。ETH处理后花楸胚胎第3天即可萌发,早于其他处理,且在第8天时萌发率达到最高。结果表明,ETH对种子萌发具有促进作用。
此外,向Na2CO3、NaHCO3中加入ETH后,花楸胚胎的萌发率又显著提升,说明ETH可以缓解Na2CO3、NaHCO3对花楸胚胎萌发的抑制,但具体的作用机理还尚不明确,需进一步进行研究。
3.3 外源ETH对盐碱胁迫下的花楸胚胎萌发有调控作用
有研究发现,外源添加ETH、赤霉素、油菜素内酯等化学物质会缓解盐碱胁迫对植物种子萌发和生长的抑制作用[24]。外源喷施ETH对盐胁迫下萌发受到抑制的种子是有效的[25]。综合ETH处理对正常情况下以及Na2CO3、NaHCO3胁迫条件下花楸树胚胎的萌发率、发芽速度、发芽指数和发芽势的结果可知,外源ETH不仅可使正常休眠的花楸胚胎萌发得更整齐、更快、萌发率更高,还对盐碱胁迫下的花楸胚胎休眠有一定的解除作用。说明在花楸树胚胎休眠解除和盐碱胁迫解除中ETH具有重要的调控作用。
综上所述,本研究发现外源ETH可以促进花楸胚胎萌发,盐碱胁迫会抑制花楸胚胎萌发,外源添加ETH可以有效地改善盐碱胁迫对花楸胚胎萌发的抑制作用。本研究结果为建立外源ETH促进花楸胚胎萌发技术提供了科学依据,并可为花楸在盐碱地区推广栽培提供参考。
猜你喜欢
热带作物学报(2022年7期)2022-08-06
舰船科学技术(2022年11期)2022-07-15
作物学报(2022年8期)2022-05-29
金桥(2021年11期)2021-11-20
东坡赤壁诗词(2018年2期)2018-05-10
现代农业研究(2017年3期)2017-04-17
江苏农业科学(2016年4期)2016-06-14
江苏农业科学(2015年11期)2016-01-27
吉林农业(2014年5期)2014-07-09
食品工业科技(2014年21期)2014-03-11
