
免灌人工梭梭林生长与土壤水分变化的耦合关系
2022-10-25 02:12朱家龙周智彬王立生姜咏雪
干旱区地理(汉文版) 2022年5期
朱家龙, 周智彬, 王立生, 吕 平, 姜咏雪
(1.中国科学院新疆生态与地理研究所国家荒漠-绿洲生态建设工程技术研究中心,新疆 乌鲁木齐 830011;2.中国科学院新疆生态与地理研究所荒漠与绿洲生态国家重点实验室,新疆 乌鲁木齐 830011;3.中国科学院新疆生态与地理研究所莫索湾沙漠研究站,新疆 石河子 832000;4.中国科学院大学,北京100049;5.新疆生产建设兵团林业和草原工作总站,新疆 乌鲁木齐 830013;6.新疆瑞绎昕生态园林技术有限公司,新疆 乌鲁木齐 830011)
梭梭(Haloxylon ammodendron)是藜科多年生灌木状小乔木,在我国主要分布于西北荒漠,具有耐干旱、耐风蚀、耐高温、耐严寒、耐瘠薄等特征[1-3]。准噶尔盆地地处干旱区,水资源匮乏,风沙危害严重,生态环境脆弱[4]。作为该沙漠中规模最大、分布最集中的物种,梭梭是准噶尔盆地绿洲边缘免受沙尘暴侵蚀的天然屏障。然而,近半个世纪以来,由于气候变化以及过度采伐、放牧、不合理开垦等人为因素,使准噶尔盆地内梭梭种群面积减少,植被景观破碎严重[5];梭梭种群幼苗正常补充机制受到严重威胁,种群年龄结构普遍呈现衰退,群落显现逆行演替特征[6];与此同时,古尔班通古特沙漠的流沙化面积不断地增加,沙漠化现象日趋严重[7],严重威胁到以梭梭为建群种或优势种的植被群落履行绿洲生态屏障功能。
自20 世纪80 年代以来,梭梭一直是古尔班通古特沙漠周边人工造林的首选树种。莫索湾地区与玛纳斯流域建立的主要由人工梭梭林及梭梭天然荒漠林组成的梭梭防护林体系被认为是沙漠地区人工生态工程成功的典范[8-10]。准噶尔盆地的冲积、湖泊平原上分布着大面积龟裂地,天然条件下种植梭梭的保存率很低,在龟裂地上更难得苗[11]。1980—1985 年黄丕振等[12]通过集水措施在莫索湾地区龟裂地上进行免灌溉造林,造林前期梭梭平均保存率91.1%,最低80%。植被得到较好恢复,风沙灾害明显减少,取得了较好的生态社会效益[13-14]。
但是,在中亚干旱区气候暖湿化趋势背景下[15-16],近40 a 来工程区气候和水文形势发生了变化,尚不清楚这种变化会对免灌溉人工梭梭林产生什么样的影响,梭梭林分生长现状如何?现状产生差异的原因是什么?针对采取集水造林措施的梭梭林地,以不同株行距建植的梭梭林植被为研究对象,布设样地,开展植被群落调查、监测、取样、分析和评估工作,阐明免灌溉集水造林条件下不同密度梭梭林的林分生长及结构特征,探讨梭梭生长与土壤水分的相互影响,为绿洲边缘地带人工梭梭林建设提供理论依据和技术支撑。
1 材料与方法
1.1 研究区概况
研究区位于古尔班通古特沙漠西南缘的莫索湾沙漠研究站(45°02′N,86°06′E;海拔346 m),地处荒漠与绿洲的过渡带区域。该区属于温带大陆性干旱气候,年均气温6.6 ℃,年均降水量不足120 mm,蒸发量高达1942 mm,地下水位13 m左右[17-18]。冬季积雪厚度约13 cm左右,最厚可达27 cm,初春(3—4月)温度回升,冻土和积雪融化,蒸发较弱,使土壤含水量增加。地带性土壤属荒漠灰钙土,部分土地发生碱化[19]。该区人工梭梭林于1984年种植,林地内无其他灌木生长。
1.2 研究材料的选取
选取采用集水措施的梭梭林地,即采用修集水沟的办法在地势平坦的龟裂土地上建植梭梭。1983年秋季开沟,两边翻土,沟深25 cm,沟距3~10 m,边坡约6°,1984年在沟底按株距1~6 m挖植树穴,穴内栽植梭梭,造林后至今38 a未进行人工灌溉[13,20]。根据各集水造林地沟距与株距的不同,本研究选取了不同造林密度的梭梭防护林体系作为研究对象。
1.3 观测样地设置
2021 年4 月,采用典型抽样法在集水造林地选择初始造林密度为株行间距1.5 m×7 m、4 m×4 m和6 m×3.5 m 的3 个不同密度且集水沟均为南北走向的梭梭标准地(基本不受人为及病虫害扰动),分别设置20 m×50 m 调查样地进行试验(图1)。在垄间及其左右两侧坡中位置设置1 组3 个1 m×1 m 的林下植被调查小样方(图2),各样地沿对角线方向分别设置3组。

图1 各林地梭梭生长状态Fig.1 Growth state of Haloxylon ammodendron in each forest land

图2 样地调查示意图Fig.2 Schematic diagram of sample plot investigation
1.4 梭梭生长状况观测
2021 年5 月,在每个20 m×50 m 样地内查数和量测所有的梭梭,调查内容包括:株高、冠幅、地径、枯枝比例、健康状况及林下植物盖度,并根据梭梭根部的精准定位及株高、冠幅、地径等指标的实地调查,将各样地内的梭梭划分为母树(初始人工种植梭梭)、幼树和幼苗(株高≤40 cm),参照丁改改等[10]I龄级的梭梭进行划分。在每个样地内利用之字法分别选取具有代表性的15株母树、15株自然更新苗(包含10株幼树和5株幼苗)作为标准样株,并进行挂旗区分,于2021年5—9月每月中旬测量株高、冠幅、地径及新枝生长量(每木15枝)。
1.5 林木生物量计算
本文选用李钢铁等[21]建立的回归模型计算梭梭单株生物量,该模型的复相关系数为0.99,拟合率为86.5%,将单株梭梭生物量累加得到整个研究林地的生物量。

式中:BW为单株梭梭的地上生物量(t·hm-2);RW为单株梭梭的地下生物量(t·hm-2);H为株高(m);E为冠幅(m2);R为地径(cm)。
1.6 土壤水分测定
在3个样地内梭梭保存率较好的区域分别选择2 株具有代表性且长势较好的母树,在其株间和其中1株梭梭根部、坡中、垄顶分别设置4根土壤水分检测管(图2),于2021 年5—9 月每月中旬利用TRIME-TDR 土壤水分测试仪读取0~280 cm 深土壤体积含水量,每隔20 cm读数1次,3组重复。
1.7 数据处理与分析
利用SPSS 20.0软件进行数据整理和分析,通过在One-Way ANOVA 在95%的置信水平上,采用Duncan法进行显著检验,所有统计分析在0.05显著性水平与0.01 极显著性水平上实现。利用Spear⁃man 相关分析梭梭株高(H)、冠幅(E)、地径(R)、地上生物量(BW)、地下生物量(RW)、总生物量(BW+RW)与垄顶、坡中、根部、株间及梭梭根部不同深度土壤水分的交互影响。运用Origin 2021绘图。
2 结果与分析
2.1 不同密度梭梭林分结构特征
2.1.1 梭梭保存与自然更新从表1 可以看出,各密度林地母树保存率(枯萎、伏地和断裂均不记在内)随造林密度减小而增大,其中低密度明显高于中密度和低密度;母树保留密度最高为中密度,最低为高密度;梭梭自然更新苗与母树数量之比可以反映种群的自然更新能力,梭梭更新苗占比均在85%以上,其中幼苗占比均在7成左右,各林地自然更新比均在3 以上,说明在长期免灌溉条件下的3种密度梭梭林依然有较好的更新能力和可持续性,造林密度越小的林地自然更新能力相比表现越好。人工林地郁闭度与林下植被盖度的大小,直接影响着固沙林分的防护效果,各林地郁闭度均大于10%[22-23],郁闭度与林下植被盖度变化趋势依次为低密度>高密度>中密度。

表1 各梭梭林地基本特征Tab.1 Basic characteristics of Haloxylon ammodendron forest land
2.1.2 梭梭生长特征
(1)整体长势与生物量
从表2 和图3 可以看出,高密度母树的平均株高、平均冠幅、平均地径均高于其他林地,变化趋势与母树生物量相同,其中中密度母树的平均株高仅为158 cm,远低于高密度和低密度;中密度自然更新苗的平均株高与平均地径均显著高于其他林地,变化趋势与自然更新苗生物量相同。各林地母树平均枯枝比无显著差异,但低密度自然更新苗的平均枯枝比要显著好于中密度林地(图3)。
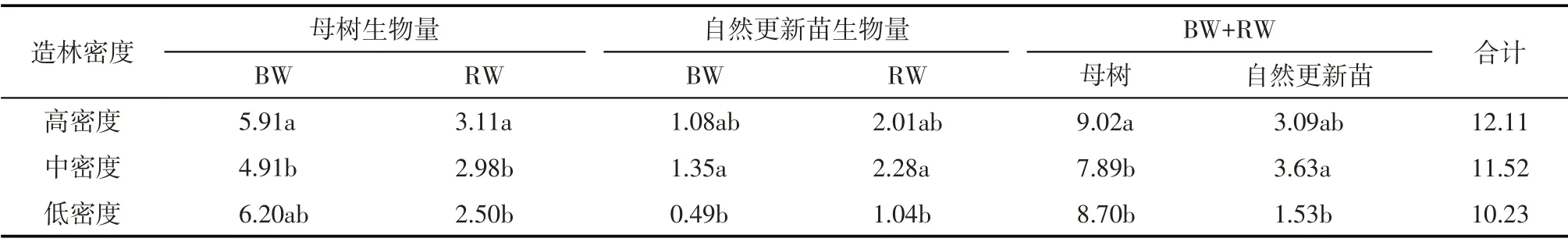
表2 梭梭林木生物量Tab.2 Biomass of Haloxylon ammodendron forest /t·hm-2

图3 各林地梭梭林木平均生长量Fig.3 Average growth of Haloxylon ammodendron in each forest land
各林地梭梭林木总生物量为10.23~12.11 t·hm-2,其中母树总生物量随母树保留密度递增呈逐渐下降趋势,依次为高密度>低密度>中密度;母树地上生物量变化趋势与林下植被盖度及梭梭郁闭度相同,依次为低密度>高密度>中密度。自然更新苗生物量为1.53~3.63 t·hm-2,其中最高值的中密度要比最低值的低密度高出2.10 t·hm-2。
(2)生长季生长特性
在各造林密度的林地内,梭梭生长季的生长速率差异不明显(图4)。母树的株高随时间变化不大(受干梢影响),平均冠幅变化趋势均表现为先在5—7月增加后在8、9月逐渐放缓,地径在5—9月呈逐渐增高趋势(高密度除外);梭梭更新苗的株高、冠幅和地径在5—9 月均呈缓慢增高的趋势,其中,低密度更新苗的株高和地径增长趋势最为明显。
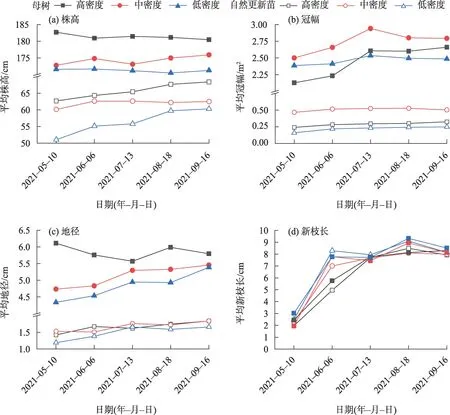
图4 各林地梭梭生长量月变化Fig.4 Monthly variation of Haloxylon ammodendron growth in each forest land
各林地内梭梭新枝长度个体间差异明显,如高密度最长达16.94 cm,最短仅为1.98 cm,但15 株母树标准株与15 株自然更新苗标准株求均值后在各月份的差别不大,整体在5—7月快速增长后在8月达到最大值,此时各林地平均枝条长度从大到小依次为低密度(9.21 cm)、中密度(8.51 cm)、高密度(8.31 cm),9月因白粉病有所降低。
2.2 不同密度梭梭林地土壤水分变化特征
2.2.1 土壤水分垂直变化从图5 可以看出,0~20 cm表层土壤的体积含水量月变异系数最大,主要受降水和短命植物的影响;高密度与低密度在20 cm以下随深度增加土壤体积含水量先趋于稳定后分别在220 cm与260 cm土层上升;中密度则呈波动变化,在60 cm、100 cm、140 cm 和180 cm 出现明显拐点。各密度林地140 cm 土层上下的体积含水量差异明显,利用多重比较发现,中密度与其他林地在140 cm土层以上的体积含水量存在极显著差异(P<0.01),高密度与其他林地在140 cm土层以下的体积含水量存在极显著差异(P<0.01),分别取平均值发现:0~140 cm 土层的体积含水量表现为中密度(10.55%)>高密度(7.46%)>低密度(6.45%);140~280 cm 土层的体积含水量表现为高密度(6.94%)>中密度(4.43%)>低密度(4.21%)。总体上在地表近300 cm深的范围内,土壤体积含水量由大到小依次为中密度(7.49%)>高密度(7.20%)>低密度(5.33%),垂直变异性表现为中密度(0.44)>低密度(0.34)>高密度(0.22)。

图5 各林地土壤水分垂直分布Fig.5 Vertical distribution of soil moisture in each forest land
2.2.2 不同位置观测点土壤水分变化实地调查显示中密度林地垄沟的边坡接近0°,降水在地表几乎不发生汇流,低密度林地沟距较窄,降水沿坑壁向四周入渗,枝叶对降水的拦截与根系对水分的吸收使得梭梭根部水分明显低于株间及垄沟其他位置,表现为垄顶>坡中>根部,株间>根部;但高密度林地沟距宽7 m,为垄间汇聚大量水分并向深层次的根系入渗,表现为根部>坡中>垄顶,根部>株间(表3、图6)。

图6 各林地不同观测点土壤水分垂直分布Fig.6 Vertical distributions of soil moisture at different observation points in each forest land
2.3 人工梭梭林生长特征与土壤水分关系
2.3.1 母树生长特征与其根部土壤水分相关性从图7 可以看出,母树生长与140 cm 土层以上土壤水分无显著相关性,与140 cm土层以下土壤水分负相关性明显;140~180 cm 土层,除地下生物量外母树的各项生长指标均与土壤水分呈显著或极显著负相关;220~280 cm土层,母树的冠幅、地径及地上生物量均与土壤水分呈显著或极显著负相关。
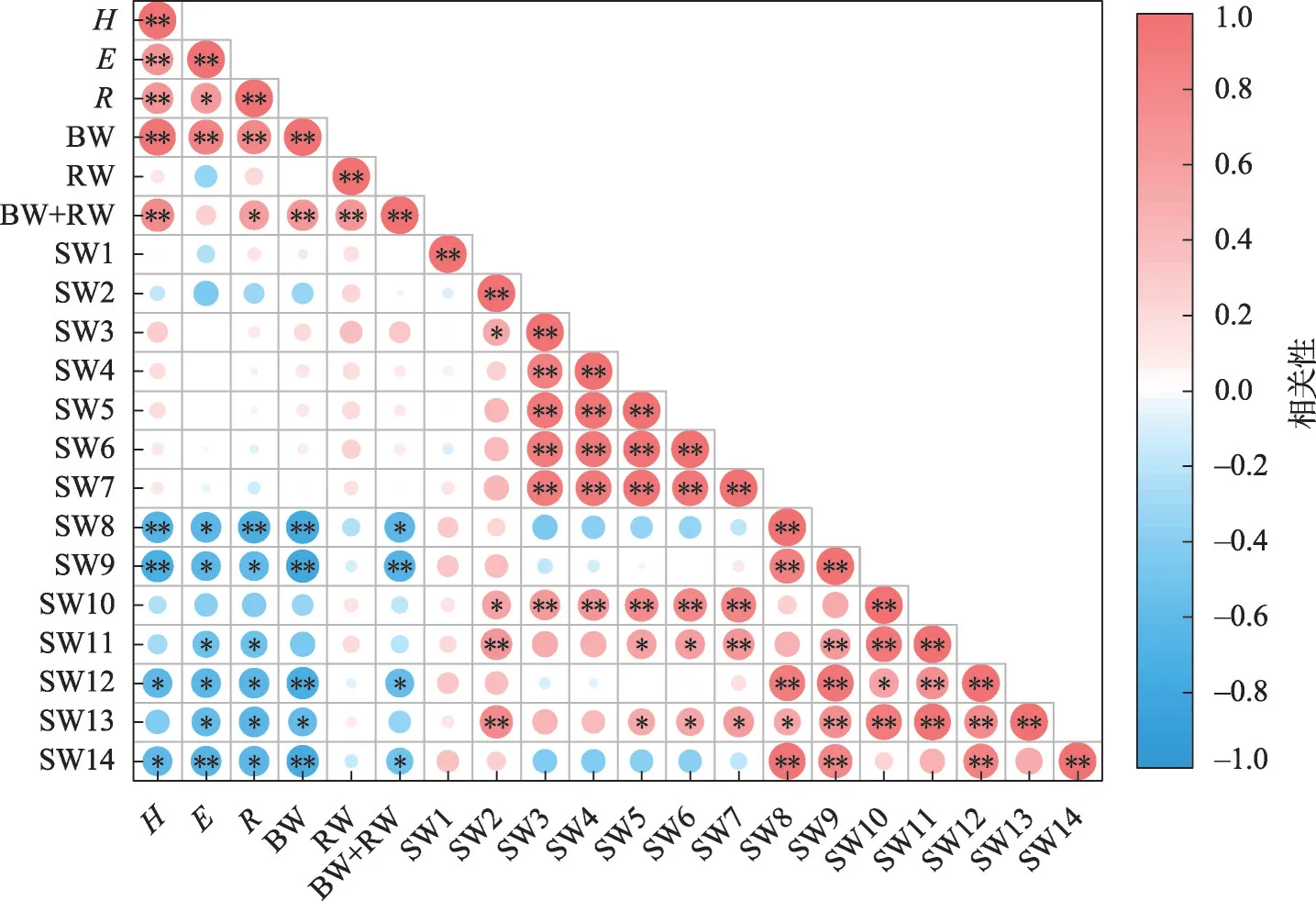
图7 母树与其根部土壤水分相关性Fig.7 Correlation between the growth of artificial Haloxylon ammodendron and its root soil moisture
2.3.2 母树生长特征与其株间土壤水分相关性从图8 可以看出,梭梭的各项生长指标与0~20 cm 土层土壤水分正相关;与60~100 cm 土层土壤水分负相关,特别是地径与该层水分呈显著负相关;与120~240 cm 土层土壤水分正相关(地下生物量除外),但不显著。
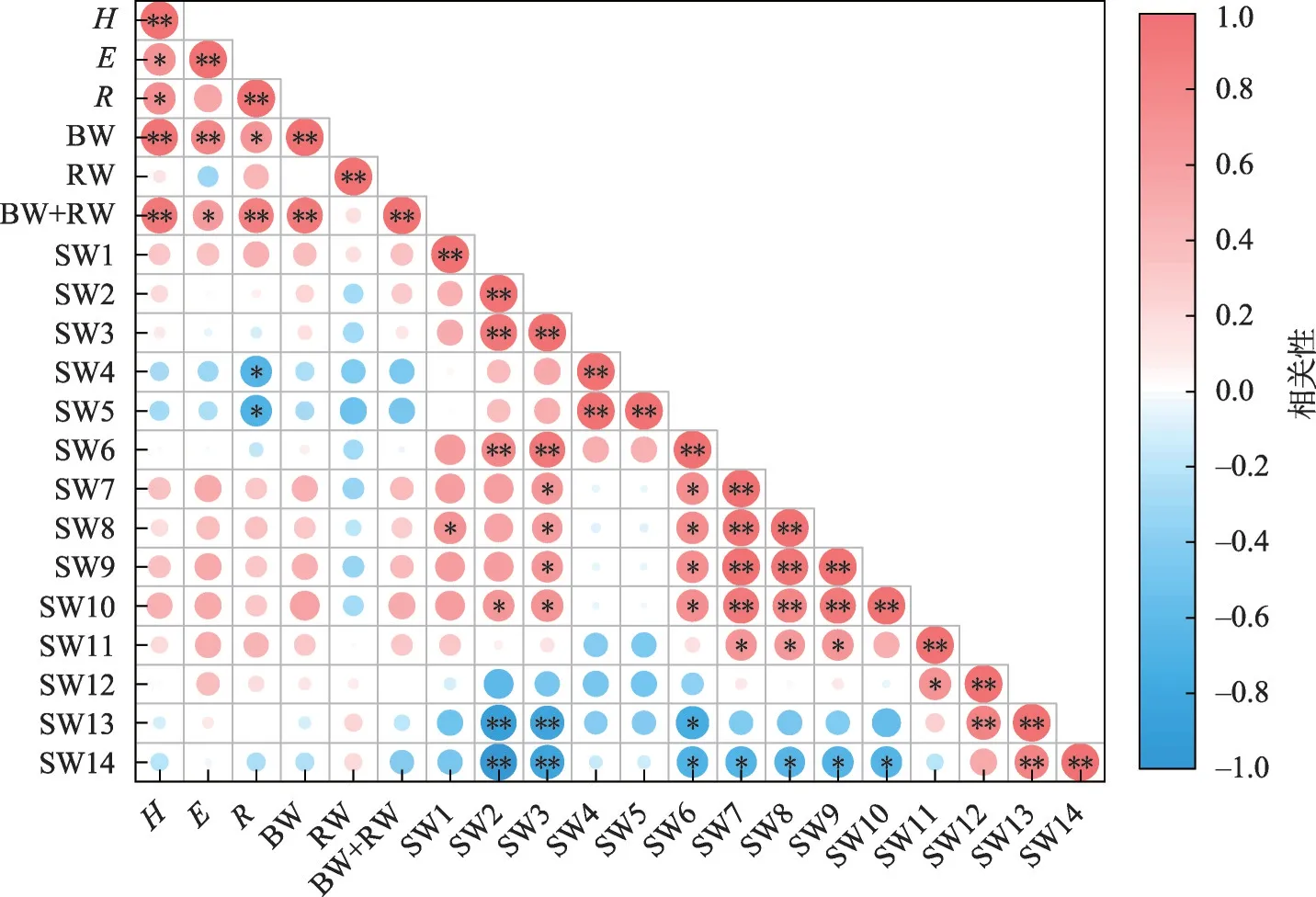
图8 母树与其株间土壤水分相关性Fig.8 Correlation between the growth of artificial Haloxylon ammodendron and soil moisture among its plants
3 讨论
3.1 不同造林密度梭梭林地土壤水分差异原因分析
各林地土壤水分的差异一方面受土壤表面微地形影响,发生移集作用使集水区的湿层加深加厚,因而土层中蒸发耗失的水分将随之显著减少,但随年限增长个别林地土壤微地形趋于平坦,降水平均铺散在地面,被湿润的土层往往较浅。研究区林地水源主要来源于初春(3—4月)融雪补给期,若不进行微地形改造,其他季节的水很难使深层土壤汇入大量水源[24]。一方面受土壤质地的影响,研究区除表层为龟裂土外,地下主要是黏土及对水的束缚性较差的沙壤土[25]。沙土干透而成隔离层,沙土层以下的黏土因受上层沙土的隔离而水分得以保留,成为沙下的贮水体。在准噶尔盆地的龟裂土,质地黏重,黏土表层寸草不生,但龟裂土的含水量与沙土层厚度密切相关[24]。中亚地区,沙土25~30 cm厚的表层土可能因蒸发而干透,但该层以下水分将保存在土体中供植物利用[26]。另一方面土壤水分的差异与梭梭根系的吸收利用有关,对比梭梭根部水分与株间水分可知,观测林地母树株高、冠幅较大,根部分布的林下植被盖度较高,降水未到地面便被蒸散发损失掉,阻碍了根部及其附近水分的补充;母树的生长发育消耗利用了梭梭根部140 cm以下土壤水分,其中对140~240 cm土壤水分的吸收更为明显;同时,由于梭梭根系呈倒“金字塔”型[27],梭梭在其主根周围广泛分布着侧根,在距主根200~300 cm 水平距离内梭梭侧根主要消耗利用60~100 cm土层深的水分,表明梭梭为补偿根部土壤水分的亏损,只有通过根系不断向深层、向四周土层延伸来吸收水分以维持体内水分平衡[28-29]。
梭梭根部附近是固定沙面,位于各林地垄间,其林下植被盖度、地表生物结皮盖度较大,对降水有显著截留作用,同时梭梭消耗根部附近水分用于生长发育,故中、低密度建植的林地主要表现为距梭梭根部愈近土壤水分愈低。高密度建植的梭梭林地则相反,表现为距梭梭根部愈远土壤水分愈低,这是由于高密度林地的垄间距(行距7 m)及垄间坡度(21°)远大于其他林地,为根植于垄间的梭梭汇流大量水分。土壤水分从垄顶向垄间(梭梭根部)逐渐增加或下降的趋势,反映出不同垄间距(行距)形成的沙垄地貌坡位以及梭梭根系对降水、径流、入渗、蒸发和蒸腾等水文过程有较大的影响作用[30]。
3.2 梭梭生长差异原因及土壤水分利用分析
梭梭生长量不仅受密度影响,还受到土壤水分的制约[31-33]。种植密度较大的人工林若不通过间伐等措施来缓解土壤水分的消耗,梭梭会逐渐死亡[34];梭梭个体会随着生长而对水分产生激烈竞争,从而导致强烈的自然稀疏[35];修筑集水沟时,一些地段将因集水面积过大,导致集水量超过梭梭生长所需而引起死亡[36]。众多研究表明,在干旱地区土壤水分是影响植被生长最关键、最直接的生态因子[37-38],本文3 种造林密度的梭梭林在长期免灌溉的条件下,土壤坡面的质量含水量(根据实测坡面平均土壤容重1.3 g·cm-3换算)均在2%以上,能够满足梭梭的长期生长发育和自然更新[39]。土壤结皮与土壤表面微地形的差异也是影响梭梭生长发育的重要因素[40-41]。梭梭生长量会随林分密度的减小和集水沟汇流量的增加而增大[42]。因此,确定适当的梭梭集水造林密度,并根据林分生长过程中的变化采取重塑集水沟、保持合理株行距、间伐等措施,改变土壤水分的再分配,从而直接或间接影响梭梭的生长与自然更新[43],使林分结构保持长期稳定,更大限度地发挥绿洲边缘人工梭梭林防风固沙的效益。
本文研究表明梭梭林郁闭度、林下植被盖度越高,0~140 cm土层含水量越低,梭梭更新苗(主要利用表层土壤水分)长势越差;0~140 cm 土层土壤水分越好,自然更新苗的长势越好、生物量越高;1983—2021 年38 a 中梭梭生长主要消耗利用根部140~240 cm土壤水分,且各密度林地内梭梭根部该土层水分越高,母树的整体长势越好、生物量越高,这与前人研究结果相似[44-45]。以上结果均表明梭梭有很强的吸水性和保水性[46]。
4 结论
梭梭作为干旱区固沙造林的先锋树种,本文对荒漠绿洲边缘地区龟裂土立地条件下免灌人工梭梭林的生长及其林下土壤水分进行了研究,得出以下结论:
(1)不同密度下38 a生梭梭的保存率和生长状况差异明显,保存率随造林密度的递增而下降,保留密度越低的林地其梭梭个体长势越好。
(2)盖度、郁闭度越高,林地0~140 cm 土层土壤水分越差,梭梭更新苗长势及生物量越差。38 a中梭梭主要吸收利用根部140~240 cm土壤水分,该层水分越高的林地,其母树的长势越好、生物量越高。
(3)当造林密度为1.5 m×7 m(990 株·hm-2)时,近40 a 后母树保留密度仅为360株·hm-2,但梭梭总体生物量更高,梭梭长势较好,林下植被更为丰富,林地土壤水分条件相对较好;当造林密度为4 m×4 m时(600 株·hm-2),林地现存的梭梭密度最高,梭梭整体长势最差;当造林密度为6 m×3.5 m时(480 株·hm-2),初始种植梭梭保存率(83.33%)、自然更新比、郁闭度及林下植被盖度均高于其他林地,但自然更新苗的长势与生物量、幼树数量均低于其他密度林地。因此,在莫索湾龟裂性土地上采取集水措施进行人工林建设时,梭梭密度保持在360 株·hm-2(株行距4 m×7 m)是较为理想的选择。
猜你喜欢
哈哈画报(2022年4期)2022-04-19
山东林业科技(2021年4期)2021-08-31
科学大众(2020年17期)2020-10-27
现代园艺(2020年17期)2020-08-31
中学生天地(B版)(2019年4期)2019-05-08
国际木业(2018年3期)2018-09-10
西部论丛(2017年10期)2017-02-23
国际木业(2016年12期)2016-12-21
广西林业科学(2016年1期)2016-03-20
绿色中国(2015年6期)2015-07-03
