
独栖蜂黄缘蜾蠃的产卵行为及其与寄生胁迫的关系
2022-10-18 13:30林木青刘益帆吴琛帆束祖飞朱朝东肖治术
昆虫学报 2022年9期
林木青, 刘益帆, 吴琛帆, 束祖飞, 朱朝东,5, 肖治术,*
(1. 中国科学院动物研究所, 农业虫害鼠害综合治理研究国家重点实验室, 北京 100101; 2. 中国科学院大学生命科学学院,北京 100049; 3. 河北大学生命科学学院, 河北保定 071002; 4. 广东车八岭国家级自然保护区管理局, 广东韶关 512500;5. 中国科学院动物研究所动物进化与系统学(院)重点实验室, 北京 100101)
独栖蜂是营独居生活,并具有筑巢习性的一类野生蜂,包括蜜蜂总科(Apoidea)和胡蜂总科(Vespoidea)等部分类群(Krombein, 1967; Staabetal., 2018)。独栖蜂在农林业和自然生态系统中扮演着传粉者和天敌等重要角色,其中以胡蜂科为代表的独栖蜂则是农林业重要害虫的天敌昆虫(Kremenetal., 2007; Rezendeetal., 2014)。因其物种多样性高和营养级复杂,独栖蜂可作为重要的生物指示类群(Tscharntkeetal., 1998)。由于受到人类活动和全球变化的影响,独栖蜂的多样性和种群数量在全球范围内呈急剧减少的趋势(Sánchez-Bayo and Wyckhuys, 2019; Wagneretal., 2021)。对这些关键昆虫进行保护是当前生态学和保护生物学领域面临的一个主要任务。然而,我们对绝大部分独栖蜂的繁殖策略仍缺少认识。因此,明晰独栖蜂的繁殖策略及其影响因素是进行农林天敌昆虫保护和利用的重要前提。
独栖蜂除受遗传调节外,通常在筑巢产卵过程中通过调节其繁殖行为来权衡产卵数量和子代性别分配。然而,巢材特征、巢材可利用性和栖息地环境因子等可显著影响其筑巢产卵行为,从而影响其繁殖成功和种群动态(Trivers and Willard, 1973; Kim and Thorp, 2001; Renauldetal., 2016)。根据巢材资源的可利用性及特征,独栖蜂的产卵策略通常受到筑巢空间的限制和寄生风险的影响。首先,独栖蜂在巢址选择上存在着空间和时间的权衡。由于筑巢空间的限制,独栖蜂在某个巢穴的产卵数量、子代性别及其排列顺序可能受到较大的选择压力。因此,独栖蜂可能通过权衡巢穴内的产卵数量和子代性别结构,以提高子代的适合度。相关研究发现,独栖蜂的孵育室数量与巢穴的固有长度呈正相关(Krombein, 1967; Santosetal., 2020)。其次,由于受寄生胁迫的影响,雌蜂可通过增加单个巢穴内的产卵数量来增加子代的存活率(Buyanjargal and Abasheev, 2015),同时通过建造更多防御性的巢室结构(如增加间室和前庭的数量)(图1)来降低被寄生风险(Krombein, 1967; Seidelmann, 1999)。一些研究也发现,独栖蜂更可能将雌性子代产在巢管内端,而将雄性子代产在巢管外端(Silvaetal., 2001; Polidorietal., 2011; Shibataetal., 2020)。这表明独栖蜂的子代性别在巢内的排列次序亦可受到寄生胁迫的选择(Santonietal., 2007)。如果巢管暴露的时间越长,外端的巢室可能更容易被寄生性天敌发现而增加被寄生风险,从而导致处于巢管外端的子代被寄生风险高于内端(Buschini and Fajardo, 2010)。因此,为了保证雌性后代的有效数量和更高的存活率,独栖蜂可能将雌性子代产于巢管内端来提高其繁殖的适合度。
黄缘蜾蠃Anterhynchiumflavomarginatum隶属于膜翅目(Hymenoptera)胡蜂科(Vespidae)蜾蠃亚科(Eumeninae)(Kim, 2003),是农林业害虫的重要天敌昆虫。黄缘蜾蠃广泛分布于中国(广东、江西、福建、香港和台湾等)、蒙古、韩国、日本、越南、老挝、缅甸、巴基斯坦、马来西亚、印度尼西亚、印度和尼泊尔等国家(Kim, 2003; Pannureetal., 2016)。黄缘蜾蠃是探究独栖蜂产卵策略及其影响因素的理想研究对象。雌性黄缘蜾蠃具有筑巢习性,常利用树洞、空茎等自然巢材进行筑巢,每个巢室被一个薄的隔板所隔开,最内端的空间为空室(empty space, EB);存放猎物并产卵的巢室称为孵育室(brood cell, BC),通常在一个孵育室内仅产一枚卵;相邻孵育室之间的空隙为间室(intercalary cell, IC);最外端的空的巢室为前庭(vestibular cell, VC)(da Costa and Gonçalves, 2019)(图1: A),在孵育室中放置鳞翅目幼虫作为其子代的食物,并以泥土对巢进行封口。因此人工巢管成为研究黄缘蜾蠃等独栖蜂繁殖生态的重要技术工具(Staabetal., 2018)。黄缘蜾蠃的子代会受到寄生性天敌的影响,其常见寄生性天敌包括钩腹蜂科(Trigonalyidae)、青蜂科(Chrysididae)、麻蝇科(Sarcophagidae)、蚤蝇科(Phoridae)等类群(Staabetal., 2016)。人工巢管研究发现,栖息地破碎化会降低蜾蠃类的筑巢率(Hashimoto and Endo, 2015)。黄缘蜾蠃子代雌性比例均与巢管的长度和直径呈显著正相关(郭鹏飞等, 2018)。综上,黄缘蜾蠃是森林生态系统中食物链的关键一环,不仅分布广,且是独栖蜂的典型代表,并有一定的研究基础,因此利用黄缘蜾蠃来探究独栖蜂产卵策略及其影响因素具有重要的和普适性的生态学意义。
广东车八岭国家级自然保护区分布有南岭山脉南缘保存完整、面积较大、分布集中、且原生性较强的典型性亚热带常绿阔叶林,其植物区系为南亚热带向中亚热带过渡类型,且其生物多样性也是整个南岭过渡带的典型代表(徐燕千, 1993)。 于2018-2020年,本研究在广东车八岭国家级自然保护区及周边区域使用人工巢管技术和公里网格方案(共计100个调查网格,网格大小为1 km2)探究了黄缘蜾蠃的产卵策略及其与寄生胁迫之间的关系。通过连续采集该蜂的筑巢巢管并饲养调查,测量了每根巢管的长度、内径和巢室特征,并详细记录了每根巢管的孵育室数量以及子代在巢管里的次序、性别和被寄生情况。 本研究拟回答以下问题:(1)巢管特征(长度和内径)和寄生胁迫如何影响黄缘蜾蠃的产卵策略? (2)黄缘蜾蠃的子代性别在巢管的排列策略是否受到寄生胁迫的影响?我们预测,黄缘蜾蠃孵育室数量和非孵育室数量受到巢管长度和内径的正影响,而孵育室数量和非孵育室数量均与被寄生率呈负相关(图1: B)。由于增加潜在暴露时间,巢管外端孵育室受到的寄生压力高于内端孵育室。因此,我们预测黄缘蜾蠃在巢管内子代性别的排列模式为在内侧的孵育室倾向于产雌性卵,而在外端的孵育室倾向于产雄性卵,且其被寄生的风险也更高(图1: C)。
1 材料与方法
1.1 研究地点
研究地点位于广东车八岭国家级自然保护区(简称车八岭保护区)(24°40′29″-24°46′21″N, 114°09′04″-114°16′46″E)(图1: A)。该保护区总面积75.45 km2,地质构造属华南褶皱系,地势西北高东南低,最高峰天平架海拔1 256 m,最低处樟栋水海拔330 m。保护区平均温度19.6℃,最高温度38.4℃,最低温度-5.5℃;年降水量1 150~2 126 mm。
1.2 巢管布设及回收观测
根据车八岭保护区生物多样性公里网格监测规范,本研究在该保护区及周边区域设置了 100个公里网格(网格面积为1 km×1 km),其中保护区内有80个公里网格和保护区外20个公里网格(图1: A)(肖治术, 2019)。每个公里网格内随机布设3个巢箱位点,位点之间的间距不少于100 m,整个实验设计共计300个巢箱位点。每个巢箱由一对长为250 mm、内径为110 mm的PVC管组成,而每个PVC管用长度相同而内径不等的芦苇Phragmitesaustralis管塞满(n=64±5)。芦苇管通常具有一个节间,节间到端口的有效长度为5~240 mm不等,内径为4~16 mm不等。巢箱用铁丝固定在树枝上,距离地面约1.5 m,巢箱口朝向东南方向。巢箱设置时,每次在铁丝上涂上凡士林(Tanglefoot®)以防止蚂蚁等侵害巢箱。
所有巢箱均在2018年5月被放置到每个公里网格,并在2018年7月至2020年11月期间按每1-2月回收被蜂类利用的巢管(表1)。已筑巢的巢管被回收后,用同等长度和内径的芦苇管进行替换,确保每个巢箱处于塞满的状态。每个巢箱的经纬度、海拔、生境和回收日期等重要信息被详细记录,所有回收的巢管均被贴上标签带回实验室用于进一步的饲养调查。
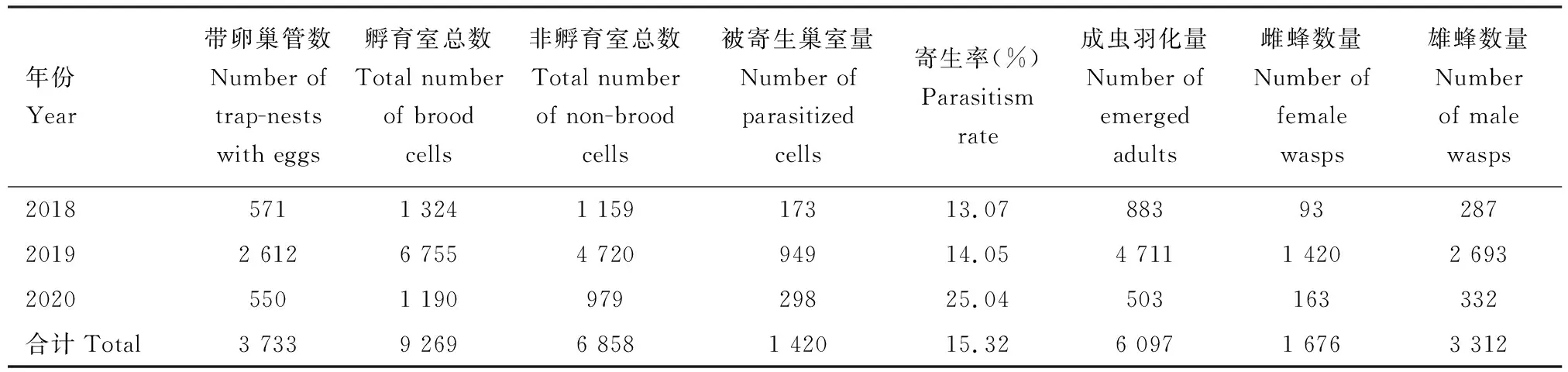
表1 广东车八岭国家级自然保护区通过人工巢管收集黄缘蜾蠃的抽样总结(2018-2020)Table 1 Sampling summary for Anterhynchium flavomarginatum using trap-nestsin the Chebaling National Nature Reserve, Guangdong during 2018-2020
在实验室内对回收的巢管进行集中处理,使用小刀纵向剖开巢管,然后测量和记录巢口的内径、巢管长度、巢室类型、寄主数量和发育阶段以及寄生性天敌类型和数量。黄缘蜾蠃的低龄幼虫通过撕咬鳞翅目幼虫吸食体液来完成发育, 老龄幼虫吞食鳞翅目幼虫头壳等坚固部位后进入预蛹期。每年8-寄生率Parasitism rate=被寄生巢室量Number of parasitized cells/孵育室总数Total number of brood cells.
12月的预蛹进入休眠期,在次年4月中下旬羽化。对已羽化的寄主和寄生性天敌均贴上标签,并用无水乙醇保存;尚未羽化的寄主连同巢管一并将放入玻璃管(规格:长度25 mm,直径6~20 mm)中用脱脂棉封口,放置在室内饲养避免阳光直晒,每隔3 d进行一次集中观察,直至巢管内所有昆虫羽化或幼虫死亡。死亡标本同样贴上标签用无水乙醇保存。成虫标本请分类学家进行了形态分类鉴定,寄生性天敌主要鉴定到目标类群,所有标本保存在中国科学院动物研究所。
1.3 数据分析
基于黄缘蜾蠃产卵的巢管特征和巢室结构,以及单根巢管的孵育室、非孵育室、子代性别和被寄生等情况,本研究综合分析了以下指标和相关因素:巢管长度为单根巢管的空室隔板到封口的距离;巢管内径为单根巢管内圆的直径(图1: A);虫室数量为巢管内孵育室,间室和前庭数量的总数;孵育室数量为在单个巢管内的产卵数量,即孵育室总数;非孵育室数量为单根巢管内所有间室和前庭数量之和;寄生率为单根巢管内被寄生的孵育室数量与孵育室数量之间的比值。
结构方程模型(structural equation modeling, SEM)通常可用来探究不同变量之间的直接和间接的关系(Shipley, 2009)。本研究使用了SEM来探究巢管特征(长度和内径)对蜾蠃虫室数量(孵育室和非孵育室)的影响,进而直接和间接地影响寄生率(图1: B)。所有变量在分析前进行均值为0、标准差为1的标准化处理。基于卡方检验的P值和相对拟合指数(cfi)来评估模型拟合的可靠程度。删除不显著的连接来完善模型,通过比较AIC值对不同模型进行筛选并选择最佳模型。使用R包lavaan(Rosseel, 2012)进行SEM分析。
根据单根巢管孵育室的数量划分为2~7组(仅2019年单根巢管内孵育室数量为2~7个,其余年份均只有2~5个,见图4)。分析中,剔除了孵育室数量少于10根巢管的组别(图2: A),每组孵育室从内到外依次计算各自的寄主雌性比例和被寄生率,计算方法为每组孵育室排序的雌蜂总量(或被寄生的巢室总量)/每组孵育室对应的产卵总量。Meta分析的方法可比较不同组别和总体情况下,子代性别和寄生性天敌数量在巢管内外的差异。首先分别构建巢管最内、外端的子代幼虫性别与寄生性天敌数量的2×2列联表,接着计算效应值和对应的研究组内方差来量化巢管最内端和最外端子代性别和寄生性天敌数量的差异。使用对数化的比值比(odds ratio)作为效应值。运用随机效应模型的meta分析对效应值进行显著性检验。Meta分析在R中用metafor包运行(Viechtbauer, 2010)。
所有的数据分析均在R 3.5.1(R core team 2018)中进行分析,并使用ggplot2软件包(Wickham, 2016)进行可视化。
2 结果
3年中,累计回收黄缘蜾蠃的巢管3 733根,孵育室总数9 269个,其中羽化成虫达6 097头,被寄生的巢室量1 420个,非孵育室总数6 858个(表1)。 黄缘蜾蠃的被寄生率平均为17.39%±5.43%,寄生性天敌以麻蝇科昆虫居多,占总寄生巢室的38%,其次是青蜂科,其占比为16%。黄缘蜾蠃羽化的子代以雄性居多(雄∶雌=1.98∶1)。黄缘蜾蠃在单个巢管内构建了2.50±1.25个孵育室(范围:1~8个),其中含1~3个孵育室的巢管占总巢管量的81%,以含2个孵育室的巢室为最多(35%)(图2: A)。 此外,单个巢管内的非孵育室数量有1.84±1.14个(范围:0~7个),其中1~3个非孵育室的巢管占总巢管量的83%,以含1个非孵育室的巢管为最多(36%)(图2: B)。
SEM分析表明,巢管长度和内径均显著正影响孵育室数量(长度:P<0.01, effect=0.47; 内径:P<0.01, effect=0.18),巢管长度也显著正影响非孵育室数量(P<0.01, effect=0.47)。孵育室数量和非孵育室数量直接显著负影响被寄生率(孵育室数量:P<0.01, effect=-0.59; 非孵育室数量:P=0.03, effect=-0.07)。巢管长度通过孵育室数量和非孵育室数量间接负影响被寄生率(total effect=-0.31),巢管内径通过孵育室数量间接负影响被寄生率(effect=-0.11)(图3; 表2)。
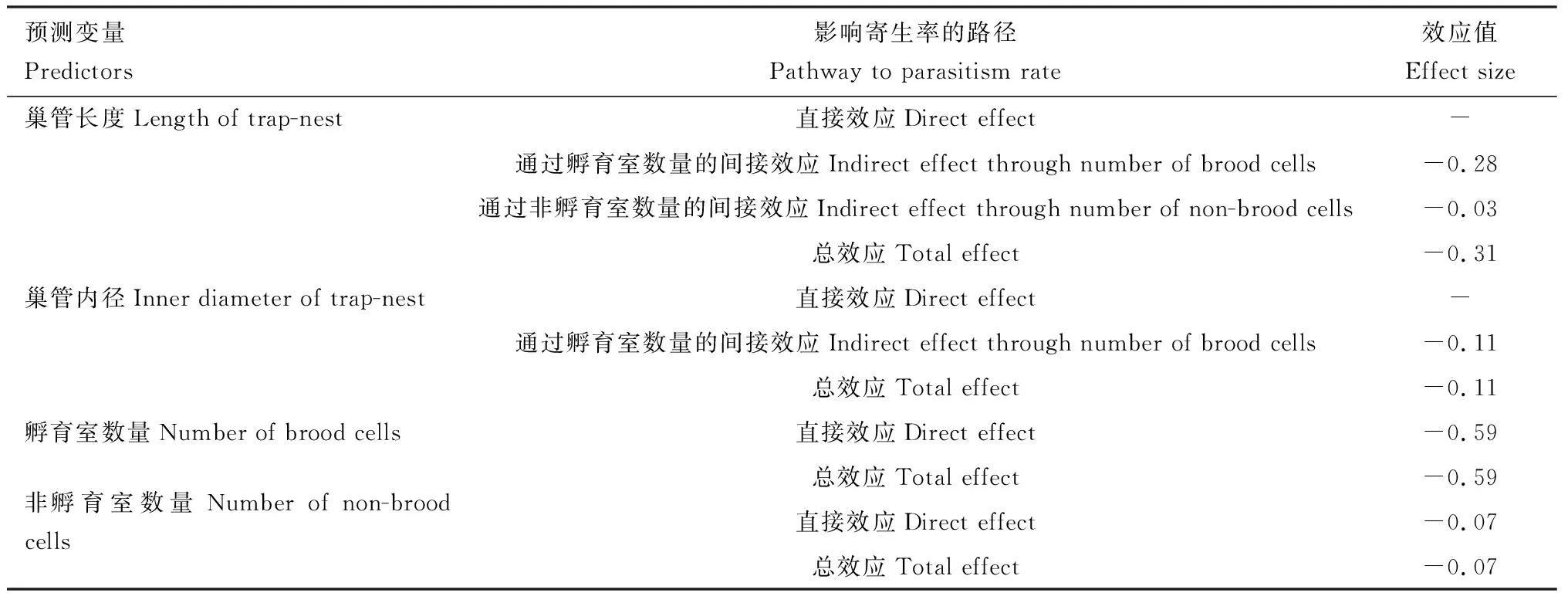
表2 基于结构方程模型的巢管特征(长度和内径)、孵育室数量和非孵育室数量对寄生率的直接、间接和总标准化效应Table 2 Direct, indirect and total standardized effects of trap-nest traits (length and inner diameter), the number ofbrood cells and the number of non-brood cells on the parasitism rate based on the structural equation models
2018-2020年孵育室数量为2-7个的巢管中,以最内端的子代雌性比例最高,而以最外端的被寄生率最高(图4)。Meta分析结果表明,巢管最内端孵育室的子代雌性数量显著高于巢管最外端的孵育室的(2018:β=1.35,z=3.83,P<0.001; 2019:β=1.94,z=5.36,P= 0.001; 2020:β=2.06,z=4.88,P<0.001)(图5: A, B, C)。2019年巢管最外端巢室的寄生性天敌数量显著高于巢管最内端巢室的(2019:β=-0.49,z=-5.02,P<0.001)(图5: E),而2018和2020年巢管最外端巢室的寄生性天敌数量亦高于巢管最内端的巢室的,但均无显著性差异(2018:β=-0.25,z=-1.24,P=0.21; 2020:β=-0.17,z=-1.05,P= 0.29)(图5: D, F)。
3 讨论
通过SEM分析,本研究发现巢管内径和长度显著地正影响黄缘蜾蠃孵育室数量,而孵育室数量和非孵育室数量显著地负影响被寄生率(图3)。在巢管最内端的子代雌性比例显著高于巢管最外端的,而最内端的被寄生率显著低于最外端的(图5)。
黄缘蜾蠃的产卵策略受到巢管特征和寄生胁迫的影响。在本研究中,巢管内径对孵育室数量具有正效应影响(图3),其可能的原因是本研究所采用的人工巢管内径(4~16 mm)处于被雌蜂所选择的范围。通常,雌蜂的体型决定了其选择巢管内径的范围,内径过小则雌蜂无法进入,降低了巢管的可利用性,内径过大则增加雌蜂搬运筑巢材料的能量,也不利于雌蜂产卵(Alonsoetal., 2012)。此外,雌蜂通过选择较长的巢管来筑巢并产更多的子代和建造更多的非孵育室(前庭和间室),可显著减少寻找其他筑巢场所所耗费的能量和被捕食的风险(Tepedino and Parker, 1984; Wearing and Harris, 1999)。同时,在寄生胁迫的环境下,黄缘蜾蠃可通过多繁衍子代和更多建造间室和前庭,以减少和抵御寄生风险来提高繁殖适合度。该结果与Buyanjargal和Abasheev(2015)对Euodynerusdantici的研究一致。研究表明间室和前庭具有抵御寄生性天敌入侵危害的功能(Coville and Griswold, 1984),而且每个巢室所形成的独立隔室也可防止相邻同类相残(Krombein, 1967)。
基于黄缘蜾蠃雌性子代和寄生性天敌在巢管内排列分布的结果,我们发现雌性黄缘蜾蠃倾向于在巢管外端产雄性子代,而在巢管内端产雌性子代。同时,寄生性天敌的寄生率在巢管最外端显著高于最内端(图5)。该类性别排列模式也出现在切叶蜂科(Raw, 1972; Hawkins, 1975)、蜜蜂科(Silvaetal., 2001; Buschini and Wolff, 2006)、蜾蠃科(Assis and Camillo, 1997; Polidorietal., 2011)、泥蜂科(Shibataetal., 2020)等独栖蜂类群中。黄缘蜾蠃的巢室由黄泥混合其唾液筑成,巢室间由隔板分隔,紧密又严实,不易破裂;黄缘蜾蠃从内到外依次建造巢室,建一个巢室产一个卵,然后即用隔板堵住。调查发现,麻蝇科、蚤蝇科和青蜂科昆虫是黄缘蜾蠃的重要寄生性天敌。这些寄生性天敌在野外环境中能通过巢入口或者在寄主外出寻觅食物的时间间隙入侵巢室(Jayasingh and Taffe, 1982; Coville and Griswold, 1984)。通常,雌蜂产卵越多,外出觅食的时间间隙越长,导致巢管暴露的时间越长,从而可能导致处于外端未封口的巢室面临更大的被寄生风险(Woginetal., 2013)。因此,独栖蜂将雌性子代更多产于内端巢室,有助于显著降低被寄生的风险,增加其繁殖的适合度(Buschini and Fajardo, 2010)。除此之外,独栖蜂的性别分配同时受到食物资源、局域交配竞争(local mate competition)等的影响(Boultonetal., 2015)。
本研究明确了巢管特征和寄生胁迫对黄缘蜾蠃产卵策略的影响,证实了寄生胁迫对该蜂子代在巢管内性别排列分布的影响。本研究通过了解黄缘蜾蠃等独栖蜂昆虫种群的繁殖策略,为进一步揭示独栖蜂与寄生性天敌之间的协同进化关系提供了新的见解,为农林天敌昆虫的保护和利用提供了科学依据。
致谢野外调查工作得到了广东车八岭国家级自然保护区管理局和当地村民的积极支持和帮助。感谢重庆师范大学的李廷景老师和中国科学院动物研究所的袁峰老师对蜾蠃及天敌的分类鉴定。感谢课题组李海东博士和欧阳芳博士在数据分析和在本文修改中提出的宝贵意见。
猜你喜欢
现代妇产科进展(2022年1期)2022-12-07
中国生育健康杂志(2022年4期)2022-11-25
生殖医学杂志(2022年10期)2022-10-19
中国药学药品知识仓库(2021年11期)2021-11-25
品牌与标准化(2021年2期)2021-07-09
中小学德育(2020年11期)2020-03-18
分析化学(2017年12期)2017-12-25
教育界·上旬(2016年12期)2017-05-25
安徽农学通报(2017年9期)2017-05-19
农业与技术(2016年22期)2017-03-07

- 昆虫学报的其它文章
- 西伯利亚蝗卵发育过程的比较转录组分析及滞育关联基因筛选
- 草地贪夜蛾烟碱型乙酰胆碱受体α6和α7亚基基因克隆及其对乙基多杀菌素胁迫的响应
- Effects of arena shape and exit location on the escapingbehavior of the Formosan subterranean termite, Coptotermesformosanus (Blattodea: Rhinotermitidae)
- 环丙氨嗪对鸡粪中亮斑扁角水虻生长的影响及添加活性炭对环丙氨嗪的解毒作用
- 多寄主型寄生蜂蝇蛹俑小蜂对新寄主的选择与适应性
- Microstructure and defense function of the acoustic organ ofPapilio xuthus (Lepidoptera: Papilionidae) pupae