
谷子矮秆突变体si-dw4的遗传分析及基因定位
2022-10-17 08:20柴晓娇白晓雷王显瑞沈轶男
江苏农业科学 2022年18期
付 颖, 柴晓娇, 白晓雷, 王显瑞, 沈轶男, 张 姼
(赤峰市农牧科学研究所,内蒙古赤峰 024031)
谷子(L.),又称为粟,脱壳后称为小米,具有自花授粉、基因组较小(约430 Mb)、抗旱耐瘠和粮草兼用等特点,是民众膳食结构改善和种植业结构调整的主体作物。近年来,随着分子生物学和基因组学研究的不断深入,谷子已经发展成为禾本科基因组研究的模式植物之一。尽管我国是谷子的起源地,拥有丰富的种质资源,同时依靠六十日、昭谷1号、矮88、吨谷等几个谷子核心资源的发现和利用,谷子育种已经取得了显著的进步,但这也造成了遗传背景狭窄,大多主栽品种仍以中秆和中高秆品种为主。因此,发掘、鉴定和利用新的谷子矮源,已成为实现谷子矮化育种最为直接的手段。此外,谷子高质量全基因组测序的完成及一些矮秆早熟品种高效遗传转化体系的建立,也为谷子功能基因组学的研究提供了扎实的基础。
我国对于谷子矮化机理的研究起歩较晚,前期主要是对谷子矮化突变体进行遗传分析和生理研究。进入21世纪之后,研究人员才陆续开始对谷子矮化基因进行定位、克隆等研究。高俊华等报道了一个谷子矮秆突变体安矮3号,并将突变基因定位在谷子3号染色体上。钱继岳通过对47份谷子矮秆材料中矮秆基因的等位性分析,发现获得的58个矮秆杂交组合中有45个组合含有非等位基因,7个组合含有等位基因。赵美丞在谷子中首次利用图位克隆的方法分离了自然矮化突变体 84133的半显性矮秆基因,其编码一个DELLA蛋白,与赤霉素(GA)信号转导有关。随后,谷子隐性矮秆基因和也相继被发现,分别定位在3号染色体和8号染色体上。近些年来,一些研究人员在谷子矮秆基因的定位、克隆方面已经取得了初步成果,但深入的基因解析尚未开展。目前,国内育种人员利用矮88及其衍生系培育出的品种大多都是中秆品种。同时很多的谷子矮秆种质还普遍存在不同程度的早衰现象,很难在育种中利用。因此,挖掘更多的农艺性状优良的矮秆资源,就成为了当前谷子矮化育种中亟待解决的问题。
谷子突变体是经过自然突变形成的一个矮秆突变体,其叶片直立,茎秆粗壮,株型紧凑,是优良的谷子矮秆种质资源。本研究以野生型 ZT002为对照,对的表型进行了系统的鉴定分析,同时利用BSA(分离群体分组分析)与QTLseq 相结合的方法对突变基因进行定位,旨在为谷子矮化育种提供可以利用的矮源,丰富和发展谷子矮化的分子机理,为进一步进行分子辅助选择育种奠定了基础。
1 材料与方法
1.1 试验材料
2012年在谷子高秆品种中发现的1株经过自然突变形成的矮秆突变体(命名为),经过多代自交纯合后,其矮化性状可稳定遗传,野生型为ZT002。
豫谷1号(Yugu1)是我国夏谷区广泛栽培的品种,并最早完成了全基因组测序。SSR41是来自韩国的一个地方品种,目前已经完成全基因组测序,该数据由中国农业科学院刁现民课题组提供。
1.2 试验方法
1.2.1 遗传分析群体及作图群体的构建 2016年夏,将矮秆突变体(母本)与株高正常的Yugu1(父本)和SSR41(父本)分别进行杂交即×Yugu1和×SSR41,该试验于赤峰市农牧科学研究所试验站(118°52′0″E、42°17′38″N)进行。2016年冬在海南三亚南滨农场种植2个杂交组合的F代,并分别收集F代真杂种的种子。2017年春在赤峰市农牧科学研究所试验站种植2个杂交组合的F代群体。2个杂交组合的F代群体均用于基因精细定位,仅有×Yugu1组合的F代群体用来做突变体的遗传分析。同时,将野生型ZT002也在同一地块进行种植,用于的表型鉴定。
1.2.2 突变体的表型鉴定及遗传分析 于植株进入成熟期时,随机选取6株和ZT002进行相关农艺性状调查。同时调查和豫谷1号、F及F群体各单株的株高,统计正常株高与矮秆植株的比率,并用卡方测验分析统计结果。
1.2.3 BSA+QTLseq法进行突变基因的定位 在2组F群体中分别选取30株极端矮秆植株,按单株剪取1~2张健康叶片用于DNA的提取,并测定其浓度,然后将30株单株的DNA 等量混合,分别构建2个矮秆混池。
通过Illumina HiSeq 2500测序平台对2个混池样本进行高通量测序,利用QTLseq法对2个混池的测序数据进行比对分析,初步确定突变基因所在的染色体和候选区间,然后在初定位的基础上利用BSA法进行基因定位。根据重测序得到的亲本之间纯合的SNP(单核苷酸多态性)和InDel(插入缺失序列)设计多态性好的标记,并利用这些标记定位基因。
2 结果与分析
2.1 突变体si-dw4的表型分析
前期研究发现,突变体叶片直立,茎秆粗壮,穗茎节显著缩短,株型紧凑,抗倒性强,是优良的谷子矮秆种质资源。在成熟期,对突变体和野生型ZT002 的茎秆部性状进行调查,发现的株高为35.90 cm,ZT002的株高为130.83 cm,突变体的株高只是野生型ZT002的27.44%(表1,图1-B)。同时发现的茎秆也更加粗壮,达到10.13 mm(表1)。对突变体与ZT002 的节间进行比对后发现,和ZT002均具有13个节间,并且每一个节间都比野生型中相应的节间短(图1-A),这与前人研究结果基本一致,都是由于节间缩短导致的矮化。

表1 突变体 si-dw4和野生型ZT002茎秆部农艺性状统计
2.2 si-dw4矮秆性状的遗传分析
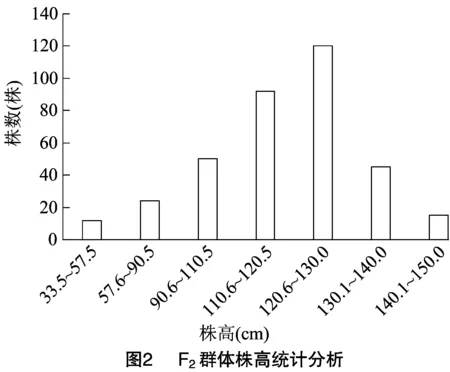
在赤峰和海南连续种植多代,均表现为矮秆性状。将突变体与具有正常株高的Yugu1做杂交,得到的F代植株全部表现为正常株高,表明矮秆性状是隐性性状。在杂交得到的F分离群体中株高分离明显,存在中间类型,通过测量358株 F群体的株高,发现最矮植株的株高为33.50 cm,最高为142.30 cm,株高位于120.6~130.0 cm区间的植株最多,且符合正态分布规律(图2)。充分说明F分离群体的株高由主效基因控制,可以把株高性状看作质量性状进行分析定位。
2.3 F2群体株高的统计分析
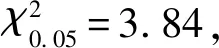

表2 F2代的株高性状分离情况统计分析

2.4 si-dw4突变基因的定位
对×Yugu1 F和×SSR41 F分离群体的矮秆隐性混池进行高通量测序。对2个混池的测序结果进行比较分析,发现SNP数量在100万~200万,InDel在20万~40万,利用QTLseq软件作图,并不能得到显著的峰值。因此,本研究利用排除法进行进一步分析,首先将×Yugu1和×SSR41 2个群体都检测到的变异位点取交集,再利用Yugu1和SSR41的全基因组数据库内的所有变异位点做过滤,结果发现一半以上变异位点集中在5号染色5~9 Mb区间内, 因此将该区间作为候选区间。根据候选区间内的InDel位点设计了多个标记(表3),并利用×Yugu1的 F分离群体进行验证,发现标记In5-6.23和In5-8.12扩增混池 DNA可以得到与母本带型一致的条带(图3),说明突变基因与这2个标记紧密连锁。根据重测序分析结果,已经几乎没有InDel位点可以使用,再无法设计出InDel标记将定位区间进一步缩小。因此,将突变基因定位于In5-6.23(6 225 727 bp)与 In5-8.12(8 123 723 bp)之间的1.89 Mb 区间内。

表3 基因定位所用的标记

3 讨论与结论
3.1 对si-dw4突变基因的分析
本研究中,对突变体和野生型ZT002的节间长度进行调查分析,发现和ZT002的节间数目一致,并没有发生变化,说明的矮化是由于节间缩短导致的。在前期对突变体农艺性状的研究中,通过与野生型相比,的叶片呈直立状,株型也更加紧凑,说明该突变基因也参与了株型的调控。前人研究表明,在水稻中dm、dn和n1这3种矮秆突变体的矮化都是节间明显缩短而导致的。同样,在玉米矮秆突变体中,最为明显的表型特征也是节间的变化,一般多表现为节间数目减少,节间长度缩短,特别是雌穗以下的节间变化尤为明显。随着研究的深入,目前在水稻、玉米、茄科和葫芦科植物中均已鉴定出这类矮秆突变体,它们的突变基因可以使节间数目减少,节间长度缩短,导致植株产生矮化,同时也发现了部分基因兼具株型调控作用,如控制叶片的直立性,使株型紧凑等。当然随着矮秆种质资源的不断利用,植物的矮化机理研究已经取得了较大进展,育成的多个矮秆、抗倒、高产、优质的作物品种也得到了迅速推广。但在谷子矮化育种中,目前可以利用的矮秆资源却仍然存在早衰和遗传基础不明确等问题,导致很难在育种中利用,阻碍了谷子的产业化进程。因而,更多新的矮秆基因的发掘不但可以为谷子矮化育种水平的提高带来更多的新突破,也为其他农作物、园艺作物的矮化改良提供了先进经验。
3.2 si-dw4突变基因的精细定位
本研究将突变基因定位在第5染色体1.89 Mb区间内,没能完成精细定位。一方面由于标记 In5-6.23 与In5-8.12之间已没有可利用的InDel位点,很难再开发出新的InDel标记;另一方面原因是作图群体数量较少,构建的遗传图谱精度较低,导致进行基因定位时检测到的重组图谱偏大而检测不到重组事件,进而表现为重组交换率偏低。为解决这些问题,本研究计划一方面基于重测序获得的SNP位点信息,利用dCAPS Finder 2.0和Primer 3(http://bioinfo.ut.ee/primer3-0.4.0/)来开发CAPS(酶切扩增多态性)标记引物,进一步加密遗传图谱;另一方面增大作图群体数量,构建精度更高的遗传图谱,精细定位突变基因。
猜你喜欢
作物学报(2022年3期)2022-01-22
河北果树(2021年4期)2021-12-02
麦类作物学报(2021年5期)2021-09-23
河北果树(2020年1期)2020-02-09
种子(2018年10期)2018-11-14
麦类作物学报(2018年9期)2018-10-09
老友(2017年7期)2017-08-22
安徽医科大学学报(2016年12期)2017-01-15
销售与市场(管理版)(2016年3期)2016-12-29
山东农业工程学院学报(2016年6期)2016-12-01
