
镉胁迫下大薸生长的变化及镉积累、分布特征
2022-10-14 07:53:36黄蕊辛建攀田如男
农业环境科学学报 2022年9期
黄蕊,辛建攀,田如男
(南京林业大学风景园林学院,南京 210037)
随着工农业的迅猛发展,生态环境中重金属(镉、铬、铜、铅、汞等)污染问题日趋严峻。重金属污染主要来源于电镀、采矿和冶炼等活动,其具有长期的隐蔽性、滞后性、累积性、区域性、不可逆性和不可降解性等特点。镉是分布最为广泛的重金属污染物之一,其在污染水体中的浓度为6~360 µmol·L,在废水中的最高浓度可以达到2 224µmol·L。镉在生物体内的半衰期长且不易降解,即使较低浓度也具有很强的生物毒性,对植物、动物及微生物构成严重威胁,甚至可通过食物链危害人类健康。因此寻找镉耐性和镉富集植物对镉污染环境进行植物修复成为当下研究的热点和难点之一。
镉被植物吸收后,会以不同形态贮藏在不同组织器官中,其在植物体内的移动性被限制,从而降低对植物的毒害作用。镉的亚细胞分布可以揭示植物对镉的耐性和解毒机制,其中细胞壁固定和液泡区室化发挥着重要作用。汤惠华等的研究表明,镉在花椰菜()叶片亚细胞中的分布表现为细胞质>细胞壁>叶绿体和线粒体。镉与植物体内的硝酸盐、氨基酸盐、有机酸盐、蛋白质、果胶酸盐、磷酸盐和草酸盐等物质结合,可以形成毒性和迁移能力不同的化学形态,这也是植物抵御镉胁迫的重要机理。WANG等研究发现镉在秋茄()根、茎、叶中以氯化钠提取态占比最高,且与醋酸提取态总占比可达到63.6%~83.6%。同时,植物还可以通过增加根系分泌物、促进重金属向衰老和脱落组织的转移来增强镉的排斥作用,以及通过增加细胞内植物螯合肽、谷胱甘肽和金属硫蛋白的生物合成和积累来增强镉的螯合作用等方式提高植物对重金属的抵御作用。
大薸()是天南星科(Araceae)大薸属()的多年生水生飘浮草本植物,具有观赏性高、吸收积累不同污染物潜力大、自然丰富度高、分布广、生长繁殖快和易打捞等优点。研究表明,大薸对镉、铬、铜、铅、锌、镍等重金属的吸收量虽不同,但其对污染水体中重金属的去除效率可以证明其具有优良的蓄积能力。同时,大薸与其他挺水植物和沉水植物的搭配种植既可满足景观需求,又可有效改善水体富营养化。除了用于净化污染水体外,大薸还可作为一种替代能源。FERNANDES等的研究表明,在进行镉污染水体的修复后,大薸还可用于生产沼气,且镉对植株所产生的甲烷活性并未造成影响。然而,由于大薸生长过于迅速,若不加以控制,则会影响当地生物多样性、危害渔业和阻碍水上交通等,将大薸应用于重金属污染水体的生态修复可在一定程度上控制其生长。因此,较低生物量和较高重金属富集量之间的平衡是保证大薸在进行重金属污染水体生态修复的同时避免因其快速生长而引起水体二次污染的关键。在镉污染水体的修复研究中,DAS等发现,大薸根部和地上部的镉富集系数最高可达2 294和870,转运系数可达0.6,对污染水体中镉的去除效果较好。SUNE等研究发现,大薸对镉的吸收主要表现为吸附、螯合和离子交换。LI等的研究表明,大薸体内的超氧化物歧化酶和过氧化物酶对镉诱导的氧化损伤具有较好的防御机制,可提高大薸对镉的耐受性。总体而言,这些研究多集中于大薸对镉的吸收富集能力以及生理适应性等方面,无法全面反映植株生长与修复效率的平衡以及其对镉胁迫的耐性解毒机制。本试验在前期预试验的基础上研究了不同浓度镉处理对大薸生物量、生长形态及其体内镉含量、镉的亚细胞分布和化学形态的影响,并探究镉胁迫下大薸生长与镉富集能力的平衡以及其耐性解毒机制,为合理应用大薸进行重金属污染水体的生态修复提供理论依据。
1 材料与方法
1.1 试验材料
试验用一年生大薸购买于江苏省宿迁市沭阳县园林绿化园艺场。试验在南京林业大学风景园林学院园林国家级实验教学示范中心温室内完成。将购买的大薸在自来水中缓苗一个月后,选取生长状况良好、形态大小一致的植株,冲洗干净后转移至盛有1/2 Hoagland营养液的透明塑料小桶(上口直径19 cm、高15 cm)中进行适应性培养,每桶2株,培养周期为15 d,期间适时补充营养液至初始体积。
1.2 试验方法
适应性培养结束后,选取生长状况良好、形态大小一致的植株进行一次性镉胁迫处理。试验采用随机区组设计,处理液由CdCl·2.5HO(分析纯)和1/2 Hoagland营养液配制而成,共设置6个镉浓度:0(对照,CK)、10(T1)、25(T2)、50(T3)、75(T4)、100(T5)µmol·L。每桶(内盛2 L处理液)2株,每个重复14株,各处理均3个重复。分别于第0、6、12、18天对植株进行形态观察和记录;于第18天时采集植株进行生物量、地上部和地下部镉含量、叶片镉的亚细胞分布及化学形态的测定,每个指标样本量为18。
1.3 测定指标及方法
1.3.1 生物量及镉含量
将植物根系置于20 mmol·LNa-EDTA溶液中浸泡15 min以去除根表面吸附的金属离子,然后用去离子水反复冲洗并用滤纸吸干。将植株分为地上部和地下部,置于烘箱中,经105℃杀青30 min后,80℃烘干至质量恒定,用电子天平称量每株植物各部分的干质量。
将烘干样品剪碎、液氮研磨并过100目筛后,称取0.1 g装入三角瓶中,加入5 mL混合酸(HNO/HClO,/=4/1)浸泡过夜,次日于电热板上220℃加热消解,直至冒浓烈白烟且溶液清澈透明,冷却后定容并过滤,采用石墨炉原子吸收光谱仪(AA900T)测定大薸地上部和地下部的镉含量。
根据测定结果计算下列指标:
转移系数=植株地上部镉含量(mg·kg)/植株地下部镉含量(mg·kg)
富集系数=植物器官中镉含量(mg·kg)/溶液中镉含量(mg·kg)
单株富集量(µg)=单株植物生物量(g)×单株植物镉含量(mg·kg)
1.3.2 镉的亚细胞分布
采用YANG等的差速离心法并略作修改。将新鲜成熟植物叶片0.2 g在预冷缓冲液[含50 mmol·LTris-HCl(pH 7.5)、250 mmol·L蔗糖和1 mmol·L二硫苏糖醇]中研磨成匀浆,4℃、3 000 r·min下离心15 min,沉淀为细胞壁组分;所得上清液在4℃、10 000 r·min下离心30 min,上清液和沉淀分别为细胞可溶性组分(主要为液泡)和细胞器组分。将各组分转移至三角瓶中,于电热板上蒸发至近干。使用与测定地上部和地下部镉含量相同的方法消煮,最后采用电感耦合等离子体质谱仪(ICP-MS,NEX I ON300)测定镉含量,同时计算各组分镉的占比。
1.3.3 镉的化学形态
采用WU等的化学试剂逐步提取法并略作修改。①称取新鲜成熟植物叶片0.5 g,加入20 mL 80%乙醇,研磨成匀浆,25℃恒温振荡22 h,8 000 r·min离心10 min,倒出上清液;再加入10 mL乙醇,相同条件振荡1 h后离心,倒出上清液。合并两次上清液,即为提取的硝酸盐、氯化物为主的无机盐以及氨基酸盐(乙醇提取态,F)。②加入10 mL蒸馏水,振荡1 h并离心,重复3次且合并上清液,提取水溶性有机酸盐等(水提取态,F)。③再加入1 mol·L氯化钠,提取果胶盐、蛋白质结合态或呈吸附态的重金属等(氯化钠提取态,F);加入2%醋酸,提取难溶的重金属磷酸盐等(醋酸提取态,F);加入0.6 mol·L盐酸,提取草酸盐等(盐酸提取态,F)。④将5次提取的上清液和残渣(F)分别蒸干和消煮后,采用ICP-MS测定镉含量,同时计算各化学形态镉的占比。
1.4 数据处理
采用SPSS 25.0统计软件对数据进行处理,对不同处理之间进行单因素方差分析和Duncan多重比较(<0.05),采用Excel 2015制图。
2 结果与分析
2.1 镉处理对大薸生物量和生长形态的影响
由图1可知,镉处理第18天时,随着镉浓度的增加,大薸地上部、地下部和总生物量均呈降低的趋势。与CK相比,经25~100 µmol·L镉处理后,大薸地上部生物量降低了27.27%~66.67%(<0.05);经10~100 µmol·L镉 处 理 后,地 下 部 生 物 量 降 低 了37.50%~56.25%(<0.05),总生物量降低了16.00%~62.00%(<0.05)。与第0天相比,镉处理第18天时,50~100µmol·L处理组大薸地上部和总生物量无显著差异(>0.05),10~100µmol·L处理组地下部生物量无显著差异(>0.05)。

图1 镉处理对大薸生物量的影响Figure 1 Effects of cadmium treatments on the biomass of P.stratiotes
由图2可知,随着处理时间的增加,大薸叶片数和分株数均呈上升的趋势;0~25 µmol·L处理组大薸冠径呈上升的趋势,而50~100 µmol·L处理组其冠径无显著差异(>0.05)。随着镉浓度的增加,大薸冠径均呈降低的趋势。第6天和第12天时,各处理组冠径与CK均无显著差异(>0.05);第18天时,50~100 µmol·L镉处理导致大薸冠径较CK降低了21.42%~29.53%(<0.05)。由图2b可知,随着镉浓度的增加大薸叶片数均呈降低的趋势。与CK相比,10µmol·L镉处理对大薸叶片数无显著影响(>0.05);第6天时,25~100µmol·L镉处理导致大薸叶片数降低了18.74%~28.12%(<0.05),第12天时降低了34.43%~52.43%(<0.05),第18天时降低了25.00%~58.00%(<0.05)。由图2c可知,随着镉浓度的增加大薸分株数均呈降低的趋势。第6天时,各处理组分株数与CK均无显著差异(>0.05);第12天时,50~100 µmol·L镉处理导致大薸分株数比CK降低了26.60%~40.00%(<0.05);第18天时,25~100 µmol·L镉处理导致大薸分株数比CK降低了28.91%~63.19%(<0.05)。

图2 镉处理对大薸生长形态的影响Figure 2 Effects of cadmium treatments on the growth form of P.stratiotes
2.2 镉处理对大薸体内镉含量的影响
由图3可知,大薸地下部镉含量和总镉含量分别为206.08~4 824.34 mg·kg和256.13~6 215.06 mg·kg,均随镉浓度的增加呈先上升后降低的趋势,在75 µmol·L时达到峰值;地上部镉含量为50.05~1 778.46 mg·kg,随着镉浓度的增加呈上升的趋势;经不同浓度镉处理18 d时,大薸地下部镉含量均显著高于地上部(<0.05),约为地上部的2.37~7.62倍。镉浓度为10~100 µmol·L时,镉在大薸地下部和总的富集系数分别为374.95~1 973.58和533.16~2 232.62,均随镉浓度的增加呈降低的趋势;镉在地上部的富集系数为158.21~399.40,随着镉浓度的增加呈先上升后降低的趋势,在25 µmol·L时达到峰值;经不同浓度镉处理18 d时,大薸地下部镉富集系数均显著高于地上部(<0.05),约为地上部的2.37~7.62倍。

图3 镉处理对大薸镉含量和富集系数的影响Figure 3 Effects of cadmium treatments on the cadmium content and bioconcentration factor in P.stratiotes
由图4可知,镉在大薸体内的转移系数为0.13~0.42。与CK相比,经10~75µmol·L镉处理18 d时,转移系数无显著变化(>0.05),而经100µmol·L镉处理18 d时,转移系数增加了75.00%(<0.05)。大薸单株富集量为128.02~1 558.61 µg,随着镉浓度的增加呈先上升后降低的趋势,25~75µmol·L镉处理显著高于其余处理组(<0.05)。

图4 镉处理对大薸镉转移系数和单株富集量的影响Figure 4 Effects of cadmium treatments on the cadmium transfer coefficient and individual enrichment of P.stratiotes
2.3 镉处理对大薸叶片中镉亚细胞分布的影响
由图5可知,大薸叶片中,细胞壁组分、可溶性组分和细胞器组分镉含量分别为0.32~154.77、0.12~85.52 mg·kg和0.39~88.89 mg·kg,均随镉浓度的增加而逐渐上升。与CK相比,经50~100µmol·L镉处理18 d时细胞壁组分镉含量增加了110.56%~482.66%(<0.05),可溶性组分增加了145.92%~711.67%(<0.05);与CK相比,经25~100µmol·L镉处理18 d时细胞器组分镉含量增加了30.74%~226.92%(<0.05)。细胞壁组分和可溶性组分镉的占比分别为34.02%~46.91%和15.15%~26.06%,均随镉浓度的增加而逐渐上升,但与CK相比无显著差异(>0.05);细胞器组分镉的占比为27.04%~50.83%,随镉浓度的增加而逐渐降低(<0.05)。当溶液中镉浓度>10µmol·L时,细胞壁组分镉的占比最大(42.61%~46.91%),其次是细胞器组分(27.04%~39.72%)和可溶性组分(17.68%~26.06%)。与CK相比,当镉浓度为25~100µmol·L时,细胞壁组分和可溶性组分镉的占比总和提高了11.11~23.79个百分点(<0.05)。

图5 镉处理对大薸叶片各亚细胞组分镉含量和占比的影响Figure 5 Effects of cadmium treatments on the cadmium content and proportion of P.Stratiotes'leaf subcellular components
2.4 镉处理对大薸叶片中镉化学形态的影响
由表1可知,大薸叶片中,乙醇、蒸馏水、氯化钠、醋酸、盐酸提取态和残渣态镉含量分别为0.02~1.10、0.12~32.35、0.40~30.67、0.91~42.39、0.06~13.98 mg·kg和0.02~0.89 mg·kg,均随溶液中镉浓度的增加呈上升趋势。与CK相比,经50~100µmol·L镉处理18 d时乙醇提取态镉含量增加了19.50~54.00倍(<0.05),蒸馏水提取态镉含量增加了82.83~268.58倍(<0.05),氯化钠提取态镉含量增加了28.78~75.68倍(<0.05),盐酸提取态镉含量增加了71.00~232.00倍(<0.05);经50、100 µmol·L镉处理18 d时残渣态镉含量分别增加了19.50、43.50倍(<0.05);经25~100µmol·L镉处理18 d时醋酸提取态镉含量增加了8.26~45.58倍(<0.05)。
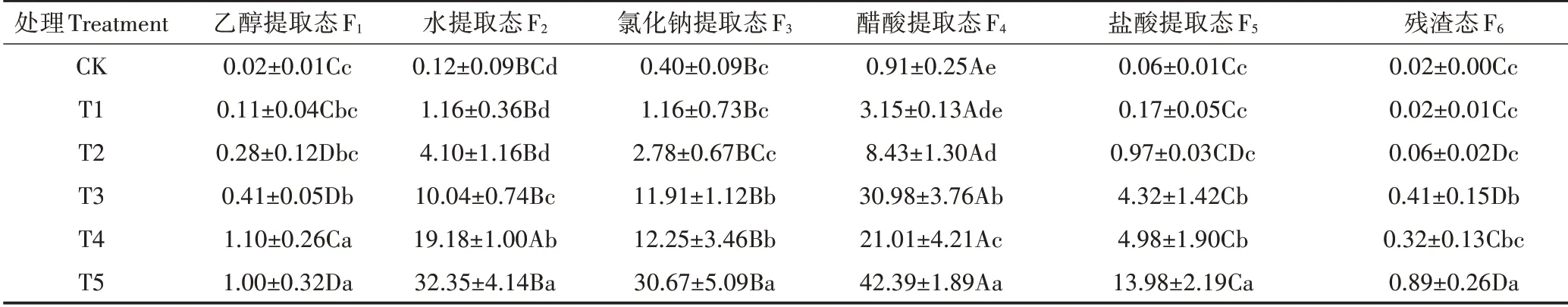
表1 镉处理对大薸叶片各提取态镉含量的影响(mg·kg-1)Table 1 Effects of cadmium treatments on the cadmium content of each extracted state in P.Stratiotes'leaves(mg·kg-1)
由图6可知,在各镉浓度水平下,均以醋酸提取态镉的占比(35.00%~59.06%)最大,其次是氯化钠提取态(16.72%~26.45%)和水提取态(7.77%~33.22%),盐酸提取态(2.92%~11.56%),乙醇提取态(0.72%~2.03%)和残渣态(0.33%~1.28%)占比较小(<0.05)。与CK相比,醋酸提取态和残渣态镉的占比随镉浓度的增加呈降低趋势,而水提取态和盐酸提取态镉的占比呈上升趋势,乙醇提取态和氯化钠提取态镉无显著变化(>0.05)。与CK相比,经10~100µmol·L镉处理后,毒性和移动性较高的乙醇、水提取态镉的占比总和提高了8.74~25.83个百分点。

图6 镉处理对大薸叶片各提取态镉占比的影响Figure 6 Effects of cadmium treatments on cadmium content proportion of each extracted state in P.Stratiotes'leaves
3 讨论
3.1 大薸生物量和生长形态的变化
根系对环境变化较为敏感,是植物接触和感应重金属的首要部位,同时也是最先受到重金属毒害的部位。本研究发现,经10~100µmol L镉处理18 d时,大薸根系生物量均显著降低(图1),表明根系受到了明显的镉毒害作用,这与谢倚慧等对马缨丹()的研究结论一致。这可能是因为多数(70.33%~88.39%)镉富集在大薸根系(图3),并通过诱发根系细胞氧化损伤、破坏根尖初生细胞DNA结构和降低其有丝分裂指数等抑制根系的生长发育。同时,环境中过高浓度的镉还会通过干扰根系对矿质营养的吸收与运输、降低抗氧化系统活性、妨碍叶片光合生理代谢等途径来影响植物地上部的生长发育。本研究发现,经不同浓度镉(≥25µmol·L)处理18 d时,大薸地上部生长被抑制,表现为生物量下降(图1)和冠径变小、叶片数减少、分株数减少(图2),这与SILVA等和COELHO等的研究结果相似。
3.2 大薸体内镉含量的变化
大薸地上部和地下部镉含量最大分别为1 778.46 mg·kg和4 824.34 mg·kg(图3),具有相当可观的镉吸收和富集能力,可作为镉污染水体的植物修复材料。转移系数是衡量植物将重金属从地下部转运到地上部能力大小的重要指标。随着镉浓度的增大,大薸转移系数逐渐上升(图4),说明镉从地下部向地上部的迁移能力增强,最大转移系数为0.42(镉浓度为100µmol·L),这与DAS等的研究结果相似。因此大薸体内镉含量的分布表现为地下部>地上部(图3),这与LI等的研究结果相似,表明大薸主要将镉富集在根系,并阻止其向地上部转运,这是大薸抵御镉毒害的机制之一。SILVA等的研究指出,12.8 mg·L镉处理会引起大薸根表皮层、外胚层以及内胚层的增厚,由于表皮负电荷的吸附作用,其厚度的增加可以加强根系作为镉生物过滤器的作用,且内胚层和外胚层可形成质外屏障,在抵御镉胁迫方面发挥重要作用,这也是大薸体内镉主要分布在根部的原因。NEDELKOSKA等的研究发现,20µg·g镉处理下,遏蓝菜()在10 d内几乎将所有吸收的镉优先分布于根毛细胞壁,然后再向共质体运输至地上部。这表明植物对镉的跨膜吸收延迟现象是其抵抗镉胁迫的重要机制,从而为细胞内重金属解毒机制的启动预留准备时间。本研究发现,大薸在镉浓度为10~100 µmol·L范围内时,地下部富集系数为374.95~1 973.58,随镉浓度增加呈逐渐降低的趋势,而地上部富集系数为158.21~399.40,呈先上升后降低的趋势(图3)。这是由于随着镉浓度的增加,大薸地上部和地下部的镉含量也增加,过多的镉离子会损伤植株根系,表现为地下部富集系数逐渐降低,而对植株地上部的损伤存在延迟效应,因此其富集系数呈先上升后降低的趋势。
10~100 µmol·L镉浓度范围内,大薸镉的单株富集量为1 051.60~1 558.61 µg(图4)。与CK相比,大薸生物量均显著降低(图1),其中25~100µmol·L处理组大薸总生物量与第0天相比无显著差异,表明在此浓度处理下,大薸既可维持较高的镉富集量,又可保持较低的生物量,在进行镉污染水体生态修复的同时还可避免因其快速生长而引起二次污染。
3.3 大薸叶片中镉亚细胞分布的变化
镉在亚细胞水平上的选择性区域分布可在一定程度上缓解其对细胞的毒害作用。细胞壁是阻止镉进入到细胞内的首要屏障,也是植物储存镉的主要部位。本研究发现,当溶液中镉浓度>10 µmol·L时,大薸叶片中有42.61%~46.91%的镉分布于细胞壁(图5),这与WANG等对苎麻()叶片镉亚细胞分布的研究结果一致。研究表明,细胞壁主要由纤维素、半纤维素、木质素和果胶等物质组成,这些物质中的羟基、羧基可与镉结合,从而降低镉离子活性和移动性。当细胞壁中阳离子结合位点达到饱和状态时,镉可经由细胞膜上的离子转运蛋白进入细胞质。镉与细胞质中的有机配体螯合后被区隔于液泡中,这决定了细胞在镉毒害过程中的生存能力。本研究发现,经10~100µmol·L镉处理18 d时,大薸叶片细胞可溶性组分中镉含量呈上升趋势(图5)。这可能印证了在较高浓度镉胁迫下,位于液泡膜上的转运蛋白可将细胞内的镉转运至液泡中,增强液泡区室化作用,从而有利于减缓镉对叶绿体、线粒体等细胞器的毒害作用。在大薸叶片中,细胞壁组分和可溶性组分中镉的占比均随镉浓度的增加而逐渐上升,其中25~100 µmol·L镉处理使其占比总和显著提高了11.11~23.79个百分点(图5)。同时,URAGUCHI等指出,镉耐性植物通常将镉固定在细胞壁,而镉非耐性植物则将镉转运至液泡。因此,大薸很可能是一种镉耐性植物,其主要通过增强叶片细胞壁固定和液泡区室化作用来应对高浓度镉胁迫,这可能也是在10~100µmol·L镉浓度范围内大薸地上部镉含量不断上升的原因(图3)。此外,本研究还发现,在镉浓度为10~100 µmol·L范围内时,大薸地上部镉含量(图3)、亚细胞分布中总镉含量(图5)和化学形态总镉含量(表1)分别为291.19~1 778.46、15.45~329.18 mg·kg和5.77~121.28 mg·kg,与10 µmol·L镉处理相比,随着镉浓度的增加,镉含量增幅分别为2.85~5.11、1.01~20.30倍和1.88~20.02倍,说明随着镉胁迫的加重,更多的镉离子被分配于植物成熟叶片中,由于成熟植物细胞的液泡相对更大,可为储存镉提供更多空间,这也是大薸应对高浓度镉胁迫的一种策略。
3.4 大薸叶片中镉化学形态的变化
镉的化学形态与其在植物体内的毒性和迁移能力有关,不同化学形态镉的毒性和活性随着提取剂(乙醇、蒸馏水、氯化钠、醋酸、盐酸)极性的增加而不断下降。唐敏等的研究表明,栾树()、臭椿()和银杏()叶片中镉主要以醋酸提取态的形式存在。在大薸叶片中,镉以醋酸提取态为主(35.00%~59.06%),氯化钠提取态(16.72%~26.45%)和水提取态(7.77%~33.22%)次之(图6),表明大薸叶片中难溶性重金属磷酸盐物质较多,由于镉主要分布于细胞壁(34.02%~46.91%),因此这些难溶性重金属磷酸盐物质可能较多分布于细胞壁中,以增强其对镉的固持能力。氯化钠提取态镉含量也与细胞壁中蛋白质、果胶等物质对镉的吸附固定作用有关,其决定了植物细胞壁对镉的解毒作用。本试验中,氯化钠提取态镉的占比仅次于醋酸提取态,这与镉主要分布于大薸叶片细胞壁有关(图5)。随着镉浓度的增加,大薸叶片中醋酸提取态和残渣态镉的占比呈降低趋势,水提取态和盐酸提取态镉的占比呈上升趋势,经10~100µmol·L镉处理后,大薸叶片中毒性和移动性较高的乙醇、水提取态镉的占比总和提高了8.74~25.83个百分点(图6)。闫雷等在黄瓜()叶片细胞镉化学形态分布的研究中也发现了相似的结果。可见在高浓度镉胁迫下,大薸叶片中的镉存在向高迁移活性转化的趋势,这说明存在于细胞壁中的难溶性重金属磷酸盐、蛋白质和果胶等物质对镉的螯合解毒作用有限。今后的研究可从分子层面揭示大薸的耐性解毒机制,即重金属亚细胞分布和化学形态相关的转运蛋白、内源信号网络的交互作用等。
4 结论
(1)镉胁迫抑制了大薸生长,表现为生物量下降、冠幅减小、叶片数量和分株数减少。
(2)大薸将大量镉积累在地下部以减少对地上部的伤害。在叶片中,细胞壁固定和液泡区室化有利于减缓镉毒害作用。镉主要以醋酸提取态存在于叶片细胞中,从而有利于降低镉的毒性和移动性。
(3)10~100µmol·L镉处理18 d后,大薸可维持较高的镉富集量和较低的生物量,在进行镉污染水体生态修复的同时还可避免因其快速生长而引起的水体二次污染。
猜你喜欢
岩石矿物学杂志(2024年1期)2024-01-29 00:15:28
中国土壤与肥料(2022年7期)2022-09-12 06:37:00
农业环境科学学报(2022年1期)2022-02-14 07:41:00
今日农业(2020年19期)2020-12-14 14:16:52
世界有色金属(2020年2期)2020-04-20 01:53:06
浙江工业大学学报(2017年5期)2018-01-22 02:03:42
广东农业科学(2017年5期)2017-08-29 10:37:54
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
食品工业科技(2014年9期)2014-03-11 18:15:40
