
珍稀濒危植物细果秤锤树群落物种组成与生态位分析
2022-10-12 06:44谭菊荣许惠春李婷婷吴初平吴正柱袁位高姚良锦
浙江农林大学学报 2022年5期
方 庆,谭菊荣,许惠春,李婷婷,吴初平,吴正柱,袁位高,姚良锦
(1. 浙江省建德市建德林场,浙江 建德 311400;2. 浙江省林业科学研究院 浙江杭州城市森林生态系统国家定位观测研究站,浙江 杭州 310023)
开展珍稀濒危植物的群落生态学研究有助于野生植物资源的保护、恢复和可持续更新。群落生态学研究一般通过探究物种的分布范围、群落结构及种内与种间联结关系等,揭示群落生活史、适应性、生长趋势等[1-3]。物种组成与群落结构在一定程度上展现植物对资源的利用能力和群落的稳定程度[4]。汪国海等[5]通过研究濒危植物单性木兰Kmeria septentrionalis的群落结构与空间分布格局,探究其聚集方式和传播途径。濒危物种的生态位宽度与群落总体关联度能够反映物种间的相互关系(竞争或促进作用)及对生境条件的适应状况和资源利用情况等[6-8]。刘万德等[9]对藤枣Eleutharrhena macrocarpa的生境特征和种间联结研究发现:藤枣与下层木呈极显著负相关,减少群落内下层木可以促进藤枣群落可持续生长[3,9-11]。杨国平等[12]通过建立预测景东翅子树Pterospermum kingtungense群落动态的Lefkovitch矩阵模型,探究濒危物种在特定的小生境片段中的分布区间。因此,基于群落生态学的研究方法,有助于全面评估珍稀濒危物种的内外致濒因子,缓解其濒危态势,实现有效的拯救保护[10-11]。
细果秤锤树Sinojackia microcarpa为中国特有的极小群落野生植物,多分布在浙江临安、建德等地,处于极度濒危和受胁迫状态[13-17]。目前,对秤锤属Sinojackia的研究相对较多。杨国栋等[18]采用生态学理论结合自组织特征映射网络(SOM)方法,划分了野生秤锤树群落的群丛类型。徐惠明等[19]分析了狭果秤锤树S. rehderiana的群落年龄结构,发现该群落具有良好的更新潜力。周赛霞等[20]研究发现:受密度制约或种子扩散限制等,狭果秤锤树的空间聚集分布趋势逐渐减弱。秤锤属物种多表现出竞争能力相对较弱,对外界干扰的响应较为显著[18-19]。本研究通过对细果秤锤树群落的长期动态监测,分析细果秤锤树群落的物种组成、生态位宽度及其与主要树种的种间关联,揭示细果秤锤树的生境适应性与竞争强度,有助于在就地、迁地保护回归实践中建立适宜的生存环境。
1 研究区概况与研究方法
1.1 研究区概况
浙江省建德市属亚热带北缘季风气候,雨量充沛,四季分明,年平均气温为17.4 ℃。土壤类型以凝灰岩发育的红壤、黄棕色壤土为主,土层浅薄且质地较为疏松,钱塘江水系中上游,境内以低山丘陵地貌为主。细果秤锤树集中分布于浙江省建德市建德林场乌石滩林区(29°32′56″~29°35′43″N,119°33′08″~119°34′05″E),主要分布在林区乌石滩、富家坞和灵山顶,海拔为23~429 m。多生长在岩石裸露率较大的山谷溪沟边的灌丛林中,呈条带状分布,群落生境数年前遭受人为砍伐干扰较严重。
1.2 样地设置与调查
细果秤锤树为典型极小群落野生植物,残存数量较少,因适存的小流域生境使得群落呈带状分布,样地设置受限。2020年8—9月,在全面踏查细果秤锤树野生群落的基础上,参照热带森林科学研究中心(CTFS)的样地建设技术规程,建立0.18 hm2的固定监测样地。使用全站仪在乌石滩、富家坞和灵山顶分别设置3个典型样方开展群落调查,共计9个10 m×20 m样方;在每个样方内设置3个5 m×5 m的下层木样地以及3个1 m×1 m的草本层样地。开展树种定位、地形测定(海拔、经纬度、坡向坡位等)、生境因子测定(土壤理化性质等)。
1.3 物种重要值计算
本研究计算上层木与下层木的物种重要值。上层木重要值=(相对多度+相对频度+相对显著度)/3;下层木重要值=(相对多度+相对频度)/2;相对多度=(某种植物的数量/样地植物的总数量)×100%;相对频度=(某种植物的频度/样地所有植物物种的频度总和)×100%;相对优势度=(某种植物的胸高断面积之和/样地所有物种的胸高断面积之和)×100%。
1.4 生态位特征与种间联结性
物种生态位特征主要采用Levins指数、Shannon-Wiener指数[21-23]反映生态位宽度,Schoener生态位相似性[24-25]与Pianka生态位重叠指数[26]反映生态相似与重叠程度。种间联结分析主要采用总体联结指数[6,8]、卡方检验(χ2)、联结系数(AC)[24]和Pearson相关系数[8,22]探究物种间关联性。采用R 4.1.0中spaa包计算生态位宽度、生态位相似性和生态位重叠程度、χ2检验、Pearson相关系数检验结果。
2 结果与分析
2.1 细果秤锤树群落野外分布与生境分析
细果秤锤树总计509株,其中富家坞分布个体数量最多(243株),灵山顶最少(71株)。群落里单丛萌蘖枝干中的最大胸径为8.10 cm,平均树高为5.40 m(表1)。乌石滩、富家坞、灵山顶细果秤锤树群落的胸径变异系数分别为34%、33%和33%,均表现为较低变异性。
细果秤锤树分布在海拔23~429 m的区域(表2和表3),乌石滩和富家坞受人工干预程度较高,存在人为滥砍及割灌除草等抚育过程。土壤呈较疏松多 孔 的 黏 质 土,土 壤 容 重 为1.06~1.19 g·cm-3,pH为4.72~5.79,偏酸性土壤,有效磷和速效钾偏低。细果秤锤树群落土壤有机质、氮、磷、钾及其速效成分中等,土壤养分条件一般。

表 1 细果秤锤树群落资源组成Table 1 Composition of population resources of S. microcarpa

表 2 细果秤锤树群落生境调查Table 2 Environmental survey of S. microcarpa population
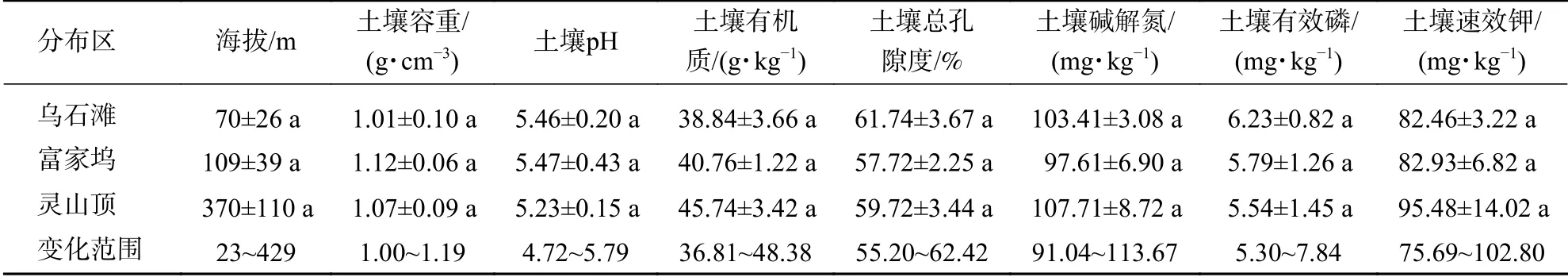
表 3 细果秤锤树群落的生境因素Table 3 Habitat factors of S. microcarpa
2.2 细果秤锤树群落物种组成
细果秤锤树样地内共记录到胸径≥1 cm的木本植物401株,隶属于35科50属51种。其中优势科有樟科Lauraceae (5属6种)、山茶科Theaceae (3属4种)、壳斗科Fagaceae (3属3种)、马鞭草科Verbenaceae (3属3种)、安息香科Styracaceae (2属3种)、大戟科Euphorbiaceae (2属2种)、金缕梅科Hamamelidaceae (2属2种)、漆树科Anacardiaceae (2属2种)、茜 草科Rubiaceae (2属2种)、榆科Ulmaceae (2属2种)。樟树的平均胸径最大,达30.8 cm,有22株;平均胸径较大的树种有臭椿Ailanthus altissima、枫香Liquidambar formosana、柏木、南酸枣和毛竹。
样地中重要值≥1%的上层木物种共16种,重要值排前4位的物种是毛竹、柏木、板栗和细果秤锤树,这4个物种重要值之和为49.85%,是群落优势树种(表4)。下层中阔叶箬竹Indocalamus latifolius的重要值最高,为15.48%;重要值排前3位的物种有水团花Adina pilulifera、毛花连蕊茶Camellia fraterna和细果秤锤树(表5)。细果秤锤树在上、下木层中重要值分别为9.50%和4.60%,是主要建群种之一。

表 4 细果秤锤树群落上层木主要物种的重要值和生态位宽度Table 4 Important values and niche breadth of the dominant species in upper wood layer of S. microcarpa community
2.3 细果秤锤树群落生态位宽度
细果秤锤树具有最大的生态位宽度,Levins的生态位宽度指数及Shannon-Wiener的生态位宽度指数在上层木中分别为5.87%和1.92%(表5),板栗、棕榈、木蜡树与柏木的生态位宽度依次降低。细果秤锤树在上层木林层与下层木林层中生态位宽度差异不明显,说明细果秤锤树的种对竞争具有一定优势,在所调查的小流域生境中具有较强的适应能力,分布幅度较广。
2.4 细果秤锤树群落生态位相似性与重叠程度
细果秤锤树群落上层木物种生态位相似性和生态位重叠值最大均为盐肤木-臭椿(表6)。细果秤锤树与上层优势树种樟树生态相似性值最高(0.62),白花泡桐次之(0.59)。生态位宽度较大的柏木和黄檀的生态位相似性达0.65,而生态位宽度较窄的枫香和臭椿的生态位相似性为0,说明生态位相似性与生态位宽度有一定关联。生态位重叠值在0.8~1.0的种对有杉木-盐肤木和南酸枣-枫香,大于0.5的种对有39对(占20.53%),其中生态位重叠值小于0.1的种对共有90对(占47.37%)。上层木树种间生态位重叠值总体偏低,对资源利用的利用策略存在差异。细果秤锤树与樟树(0.62)和黄檀(0.59)具有较大的生态位重叠,存在较大的生态和资源利用相似性。
下层木物种生态位相似性为0~0.96,生态位重叠为0~0.10,最大值种对均为海金子Pittosporum illiciodes-紫麻Oreocnide frutescens。细果秤锤树与下层优势树种檵木生态相似性值最高(0.86);与水团花(0.51)和茶Camellia sinensis(0.48)具有较大生态重叠(表7)。下层木主要物种生态位重叠平均值为0.23,且多数种对的生态位重叠在其平均值附近,表明下层木主要物种的竞争关系相对稳定。
2.5 细果秤锤树群落联结性与Pearson相关分析
细果秤锤树群落上层木12个优势种间总体联结性方差比率为1.23,大于1,即种间存在一定程度正联结;其显著检验统计量为11.05,高于χ2分布临界值,表明上层木群落间总体上呈显著的正联结关系。下层木12个优势种间总体联结性方差比率为0.58,小于1,即种间存在一定程度负联结;其显著检验统计量为5.19,介于χ2分布临界值之间,即下层木12个优势种间呈不显著负联结关系。
χ2检验主要反映不同种对之间联结的显著度。联结系数检验结果显示:上层和下层各12个优势木中,正、负联结种对数相接近。细果秤锤树群落上层木中正、负联结的种对分别为27和28个(各占种对数的40.91%和42.42%),正负关联比为0.96∶1.00。种对间总体显著率为12.12%,种间联结较松散,无联结的种对占16.67%,细果秤锤树与其他种之间都不存在联结性。下层木种对联结显著度的分布大致与上层木相似,正负关联比0.83∶1.00。细果秤锤树与水团花呈显著正联结关系。细果秤锤树-阔叶箬竹、细果秤锤树-茶、细果秤锤树-檵木、细果秤锤树-窄基红褐柃表现出极显著负关联(表8)。

表 5 细果秤锤树群落下层木主要物种的重要值和生态位宽度值Table 5 Important value and niche breadth of the dominant species in lower wood layer of S. microcarpa community

表 6 细果秤锤树群落上层木主要优势种间的生态位相似性比例和生态位重叠指数Table 6 Niche similarity and niche overlap of dominant plant species in S. microcarpa community in the upper wood layer

表 7 细果秤锤树群落下层木主要优势种间的生态位相似性比例和生态位重叠指数Table 7 Niche similarity and niche overlap of dominant plant species in S.microcarpa community in the lower wood layer
上层木中总体显著率为19.70%(极显著正关联13个,P<0.01),不显著(P>0.05)正关联25个,占37.88%;不显著负关联28个,占比42.42%。细果秤锤树与其他树种为无联结关系,整个细果秤锤树群落处于优势发展趋势(表8)。下层木中总体显著率为0,不显著正关联31个,占46.97%;不显著负关联35个,占53.03%。细果秤锤树与水团花、毛花连蕊茶、杭州榆、短柄枹栎呈不显著正关联,与阔叶箬竹、茶、檵木、窄基红褐柃呈不显著负关联。

表 8 细果秤锤树群落12个优势种χ2检验、联结系数(AC)及Pearson相关检验结果Table 8 Result of χ2 test, association coefficient (AC) and Pearson correlation coefficient of the 12 dominant species in S. microcarpa community
3 讨论
3.1 物种组成与群落结构
建德市野生细果秤锤树群落动态监测样地内树种组成相对简单,细果秤锤树多生长在次生常绿阔叶林和针阔混交林中,群落优势树种主要为毛竹、柏木、板栗和细果秤锤树。这与秤锤属调查样地内的物种组成及数量相类似[13,15-16]。调查发现:细果秤锤树群落中缺乏小径级个体或幼苗,这可能是因为秤锤属的种子萌发困难或遭受了人为的抚育等干扰,影响了幼苗的更新[13-14]。细果秤锤树是小流域生境群落中的优势种,早期生长喜较为荫蔽的环境,群落中高大上层木树种如樟树、毛竹、柏木等可在其幼苗更新时期起到遮光作用,以保护幼苗不受高温、强光照影响。在细果秤锤树生长后期,对光照需求增强,可间伐上层木,对高度接近细果秤锤树的树种进行一定程度的抚育,降低群落郁闭度[12,16-17]。
3.2 生态位宽度与生态位重叠程度
生态位宽度作为植物群落的环境适应力和资源利用能力的衡量性指标,值越大,反映物种适应能力越强,在群落中更具优势[22,27]。细果秤锤树在群落物种中重要值排在第4位,但生态位宽度却排在首位。可能是其喜光、耐贫瘠、喜微酸性土壤等生长特性有利于细果秤锤树在小溪流水域附近广泛分布。细果秤锤树的生态位宽度较大还可能与本研究的样地设置有关。本研究以细果秤锤树生长的位置为核心展开设置并调查,且呈聚集分布均匀的群落使得其占较大资源位或较大资源量,与极小群落植物圆叶玉兰Magnolia sinensis[28]、小花木兰Oyama sieboldii[29]、缙云秋海棠Begonia jinyunensis[30]在所处群落中生态位宽度均较大这一研究结果相同,表明在该分布点的研究区域生境条件下,生态位宽度大小与细果秤锤树致濒机制无必然联系。研究中有一些物种的生态位宽度大小排序与其重要值大小排序不同,如樟树、南酸枣等,这说明生态位宽度和重要值在物种之间的表现方式略有不同且并无显著关联性。
生态位相似性特征反映种间资源利用的相似程度,重叠值特征衡量生态位相似的树种在特定空间环境下资源利用的差异性,两者结合衡量种间资源竞争程度[31-33]。细果秤锤树与上层优势树种樟树和黄檀的生态相似性与生态重叠性均最高。可能是因为樟树、黄檀是对环境适应性广泛的泛化种,也可能是适合调查区域环境的特化种,因此出现与细果秤锤树较高的生态位重叠值,也表明这些种对间生态学特性比较一致,或者对生境的要求比较相似[8]。一般来说,当多个物种同时具有较大的生态位宽度时,它们之间存在较高生态位重叠的可能性更大[21]。但是,具有较大生态位宽度的物种也可能与较小生态位宽度的物种间存在较大的生态位重叠[21,31]。这是因为细果秤锤树与水团花、毛花连蕊茶为中生植物,在资源有限的条件下,它们对资源环境的竞争比较大,且对资源的利用和需求相近[32],因此,它们之间的联系也更为紧密,具有较高的生态位重叠[22,26]。且细果秤锤树所在群落中物种之间的生态位重叠程度总体偏低,说明细果秤锤树群落中大多物种对资源利用的相似程度降低,物种之间竞争较弱,生态位可通过产生分化来降低种间竞争使得物种间在群落的结构与功能上互补且稳定[7,22]。本研究发现:细果秤锤树群落大部分种对间的相关性比较弱,表明物种联结性较弱。种间负联结关系占主导,但大部分优势种种对间关联性比较低,说明样地中的不同物种间不存在紧密的相互关系,缺乏竞争或相互促进的趋势,物种间具有独立性,受外界的干扰较小[30]。
4 结论
细果秤锤树群落中物种组成较为简单,群落结构相对单一,细果秤锤树群落幼树较少,更新相对较差。细果秤锤树生态位宽度最大,在时空上占据着优势地位,属于稍耐阴、耐贫瘠、适应力较强的植物,能更好利用资源和空间。调查样地中多数树种生态位重叠度较高,大部分物种间的竞争较强,对资源利用的相似程度高。树种间不存在较显著的种间相关联结,植物种间缺乏较强的相互依赖或竞争趋势。本研究明确了细果秤锤树生存的独特环境结构和群落间相互关系,对维持其野生群落的幼苗更新和群落规模增长具有重要作用。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
贵州畜牧兽医(2022年3期)2022-06-28
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
现代园艺(2021年23期)2021-12-01
新农业(2020年18期)2021-01-07
现代园艺(2020年19期)2020-10-02
热带农业科学(2019年8期)2019-09-19
读者·校园版(2019年3期)2019-01-28
少年文艺·开心阅读作文(2017年1期)2017-02-24
