
西藏飞蝗的研究现状及展望
2022-09-28 12:19王向向顾亚欣任丽娜母泽祺唐晓琴
广西农学报 2022年3期
王向向 顾亚欣 任丽娜 母泽祺 唐晓琴, 3*
(1. 西藏农牧学院植物科学学院, 西藏 林芝 860000;2. 西藏高原资源昆虫与应用昆虫实验室, 西藏 林芝 860000; 3. 西藏高原森林生态教育部重点实验室, 西藏 林芝 860000)
西藏飞蝗(Locusta migratoria tibetensis Chen)属于直翅目(Orthoptera)蝗亚目(Locustodea)飞蝗科(Locusta migratoria)飞蝗属(Locusta), 是1963年陈永林先生在西藏自治区发现并命名的飞蝗新亚种。西藏飞蝗活动范围主要在我国的西藏自治区、青海省、四川省等高原地区, 取食广泛且食量大, 主要取食禾本科与莎草科的作物和牧草, 是一种高原重要害虫, 具有繁殖能力强、密度大、为害面积广和为害程度严重的特点。根据李婧研究发现, 在我国历代古籍中有许多处蝗灾的记载, 严重时导致庄稼颗粒无收和草场的寸草不生, 造成饥荒和出现难民, 引起社会混乱。吴志刚和秦萌研究西藏飞蝗对青稞产业的经济损失进行评估时, 结果显示西藏飞蝗每年造成的潜在经济损失达0.713亿~4.542亿元。随着全球气候变暖, 青藏高原的生态环境也受到严重影响, 导致西藏飞蝗的繁殖和发育能力增强并且增加了越冬存活率, 使西藏飞蝗暴发更加容易, 为害更加严重。本文主要对西藏飞蝗的部分特征以及综合防治措施等研究现状进行综述。
1 西藏飞蝗的形态学特征
西藏飞蝗的生长发育类型属于渐变态发育。与其他渐变态发育昆虫相似, 一生需要经历3个发育阶段。其中蝗蝻需要经过5次蜕皮、羽化才能完全发育为成虫, 即一只西藏飞蝗一生共经历5个龄期。观察发现西藏飞蝗的受精卵形状类似长椭圆形且每一个卵囊中卵的颗粒数不相等, 各龄虫态的前胸背板后缘的形状、翅芽的长度、触角的节数和体色等也各不相同, 西藏飞蝗成虫的体长、体色也会因雌雄不同有所差别。
西藏飞蝗成虫分为头部、胸部和腹部三个部分, 其头部的额唇基区有1对丝状触角, 颅侧区有1对复眼和3个单眼, 颊下区有1个咀嚼式口器。胸部的三个节分别有1对足, 共有3对足, 其前足、中足为爬行足, 后足为跳跃足;胸部含有前后2对通明翅, 前翅狭长, 具有斑纹, 后翅平常时呈折起状态, 覆盖前翅下;迁徙时, 展开进行平衡身体。成虫腹部第1节具有鼓膜器, 能发出声音, 成虫末端含有生殖器官, 因雌雄性别不同, 其生殖器的形态特征也不相同。
2 西藏飞蝗的生物学特性
2.1 西藏飞蝗的生活史
西藏飞蝗以虫卵的形式在土壤的表皮中越冬。西藏飞蝗的年生活史以四川甘孜州乡城县调查为 例, 如表1所示。由表1可知, 西藏飞蝗在四川部分地区第一年发生1~2代, 卵在土壤中越冬并于第二年3月开始孵化, 到4月为卵的孵化盛期, 5月和6月基本是以蝗蝻为主, 7月的蝗蝻开始羽化, 7月至8月为4龄蝗蝻的羽化盛期, 8月至9月为成虫的产卵盛期。西藏飞蝗的活动范围受外界温度和光照的影响比较大, 其全世代发育的有效积温相对偏高。当温度过高和过低时都会影响其取食、蜕皮、羽化等活动;成虫在光照不足或者温度较低的场所会使其产下的卵孵化率大幅度降低。
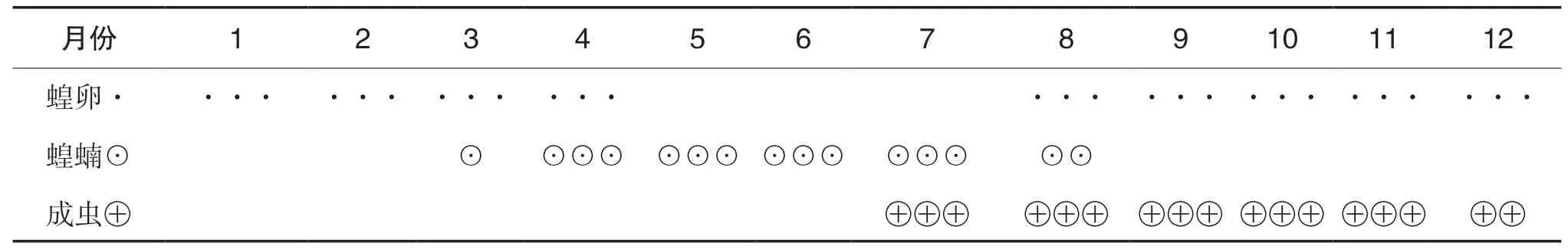
表1 西藏飞蝗的年生活史(四川甘孜州乡城县)
王思忠等利用培养箱在温度不同, 其他条件相同的条件下饲养西藏飞蝗。测量虫卵和各龄虫态的发育起始温度和有效积温, 结果显示:蝗卵、各阶段的蝗蝻、成虫的发育起点温度和有效积温各不相同。由此, 可以说明各龄虫态发育历经的时间也不相同。
2.2 西藏飞蝗的习性
西藏飞蝗成虫产卵根据地形、土壤含水量和土壤理化性质等不同有着明显的选择性。李庆等研究表明西藏飞蝗会选择坚实平坦、湿度合适的土壤产卵。
西藏飞蝗主要取食禾本科农作物和蒿草、披碱草等禾本科与莎草科杂草;在不同的寄主植物下, 其会优先取食含水量和营养价值高的植物。1~3龄蝗蝻喜欢取食幼嫩的禾本科杂草, 但是取食量相对较少;成虫期会进入暴食期, 而因青稞和小麦的长势和含营养含量此时比杂草要好, 成虫将主要以青稞和小麦为食, 造成农作物的减产。有研究证明8月的成虫密度大且是产卵的盛期, 能够在禾本科植物上获得生长发育所必需的营养物质, 但在沙棘和十字花科白菜上则不能完成整个生活史。
西藏飞蝗具有群居型和散居型两种生活类型, 但两种类型可以随其生存环境与其种群密度的变化相互转化;两种生活方式类型下的蝗虫体色、体型的大小、身体各个部分等形态学特征都有所不 同。封传红等研究证明西藏飞蝗具有群集迁徙的习性, 当西藏飞蝗虫口密度达到一定数值时, 将会由分散型转变为群居型进行迁徙。此外, 西藏飞蝗群集会受到取食的食物、虫口密度和其他行为反应的影响。陈湖海等研究西藏飞蝗触角的意义发现西藏飞蝗能够通过群集行为增加迁徙距离、增加远距离食物取食的概率。李彝利通过扫描电镜对西藏飞蝗触角外部形态结构特征观察发现不同的西藏飞蝗具有不同的感受器, 共计有5种感受器类型。感受器因蝗虫的雌雄、散居型、群居型的类型并无明显差异, 仅在数量上不同。不同虫态的西藏飞蝗对同一种人工合成化合物的触角电位反应(EAG)总体呈现散居型强于群居型、成虫强于蝗蝻、雄性强于雌性的趋势。王海建等通过实验对西藏飞蝗粪粗提物中化学聚集信息素进行测量分析, 结果发现西藏飞蝗两种生活方式下的西藏飞蝗聚集信息素组成成分相一致但所含比例有所不同。
3 西藏飞蝗的生态学特征
西藏飞蝗具有很强的迁徙能力, 以性成熟前的迁徙能力最强。唐昭华等研究西藏飞蝗成虫的生物生态学发现成虫的生殖活动、飞行能力、交尾、产卵等特性受到周围生态环境的影响。昆虫在长途迁徙过程中需要高营养的食物来维持身体所需消耗的能量, 高质量的食物会增加其成虫的大小和繁殖能力。因此, 可以改变周围的环境减少西藏飞蝗的活动范围以降低为害程度。
3.1 西藏飞蝗的耐寒性
西藏飞蝗属于变温昆虫。白天随着温度的升高其活动也频繁, 夜晚随着温度降低其活动减缓。西藏飞蝗与其他昆虫一样体内存在一种过冷却的生理现象, 西藏飞蝗的耐寒能力与过冷却点呈负相关, 西藏飞蝗虫卵的过冷却点要比其他虫态低, 即虫卵耐寒能力更强, 更容易越冬。研究发现不同发育阶段西藏飞蝗过冷却点、结冰点如表2所示, 并发现在当地冬季西藏飞蝗的存在形式只有虫卵一种。李庆等在测定西藏飞蝗的耐寒性理化指标时, 发现过冷却点和结冰点不仅各发育阶段的飞蝗之间存在差异, 而且群居型和散居型、雌雄之间也不相同。
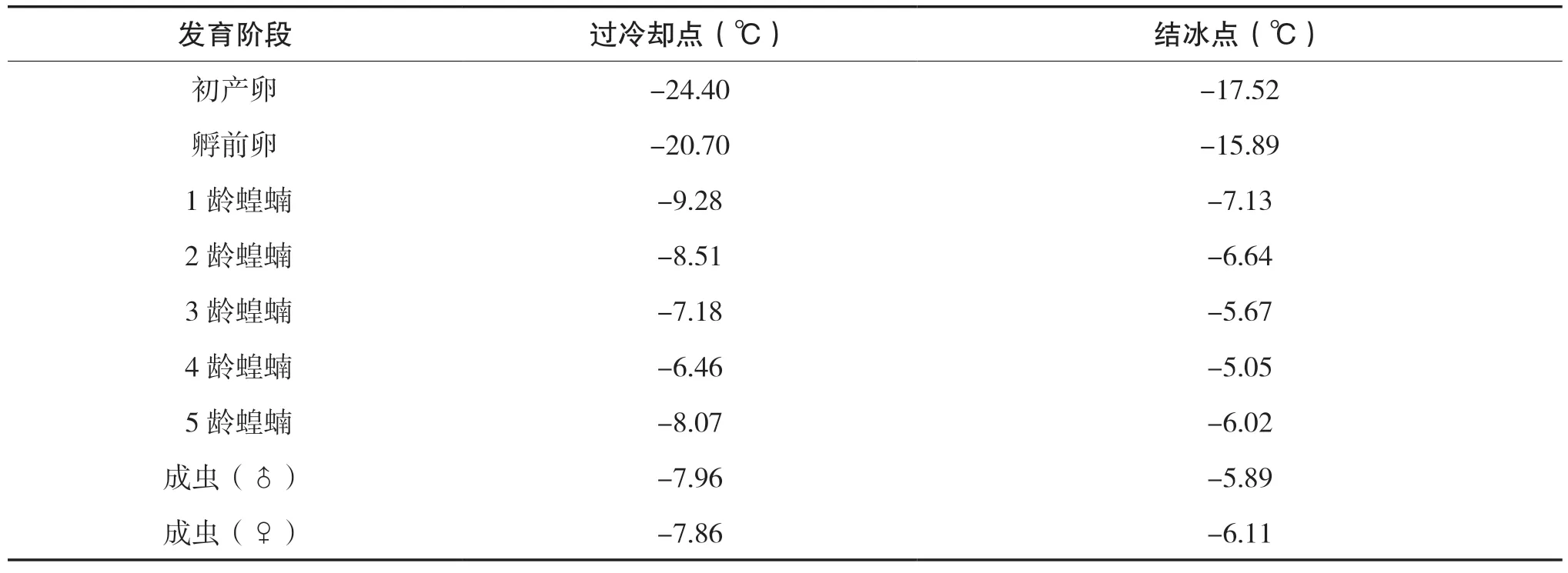
表2 不同发育阶段西藏飞蝗过冷却点、结冰点
3.2 西藏飞蝗耐寒性的理化物质
西藏飞蝗的耐寒能力与其体内的生理生化物质的含量有密切关系, 各发育阶段体内的理化物质含量如表3所示, 研究证明昆虫的过冷却点随着体内的含水量的增加而降低, 随着甘油的增加而增 加。西藏飞蝗体内抗寒性系统是以甘油等物质组成的, 为适应干燥缺氧的高原环境。在低温胁迫的条件下, 其成虫体内脂肪转化为甘油, 糖原分解成糖类小分子碳水化合物, 体内的大分子蛋白质被降解成丙氨酸等氨基酸。西藏飞蝗体内的物质通过氧化分解释放能量, 用来抵御低温的影响, 维持生长发育、生理代谢所需要的正常温度。
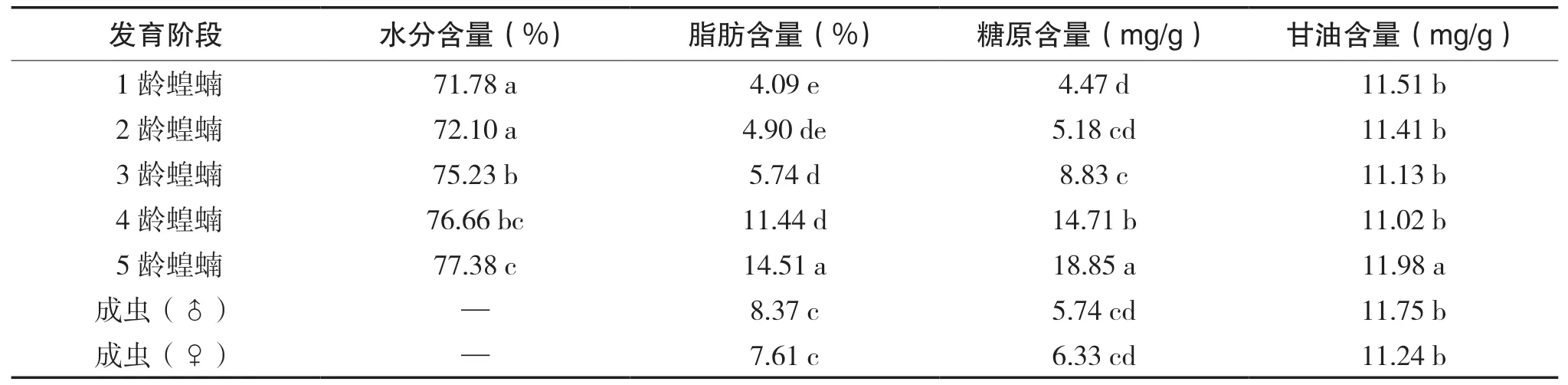
表3 西藏飞蝗(群居型)各发育阶段体内的理化物质含量
4 西藏飞蝗受环境因子的影响
当前, 全球气候还在不断地升高, 未来全球的地表温度也将继续增加, 青藏地区作为气候敏感区, 也会随全球气候的变化而变化。气候的变化将会造成生态系统的不可逆的改变, 影响着西藏飞蝗的分布。由于温度的升高导致西藏飞蝗代谢和繁衍能力都有所增强, 为害也随之加剧。董兆克等分析了全球气候变暖和局部气候恶化不仅使昆虫生存能力受到影响, 还能改变昆虫的分布范围, 是影响农作物和牧草等植物的病虫害发生的重要原因之一。
4.1 西藏飞蝗受温度的影响
西藏飞蝗的分布区域广泛且海拔差异显著, 是通过长期的适应和物种进化建立的新亚种群。温度的变化将影响西藏飞蝗生长发育的改变, 有研究证明在一定的温度范围内, 西藏飞蝗各龄期的发育历期与温度呈负相关, 发育速率与温度呈正相关, 具体见图1 。在不同的温度条件下, 其生长发育的全世代历期会随着温度的升高而缩短, 说明温度的升高将会加快西藏飞蝗的发育, 具体见图2。吴雷等研究环境因子对西藏飞蝗产卵的影响, 发现西藏飞蝗体内的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的活性在低温胁迫和高温胁迫下会发生变化来应对低温和高温的环境影响;同时, 温度的胁迫还会对丙二醛(MDA)含量影响进而影响生理代谢。韩政等分析了热激蛋白与昆虫的耐热性关系时证实热休克蛋白(HSP)是在不良环境下产生的应急蛋白, 控制应急蛋白的基因在个体受到不良环境的刺激后会转录表达, 从而提高昆虫的抗逆性。通过对比西藏飞蝗在不同的温度胁迫下的基因表达情况, 发现高温胁迫和低温胁迫条件下体内的转录基因组的表达量不同;低温下的西藏飞蝗体内的几丁质酶(CHT)活性更强, 从而提高西藏飞蝗的抗逆性能力。
图1 西藏飞蝗各虫态在不同温度条件下的发育历期
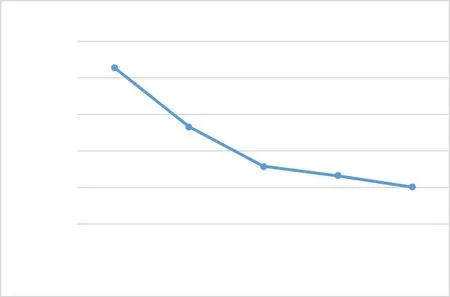
图2 西藏飞蝗在不同温度下的全世代历期
4.2 西藏飞蝗受光照的影响
吴蕾用西藏飞蝗做实验, 通过不同的紫外线辐射研究光照对西藏飞蝗的分布、分化和扩散, 发现用不同紫外线辐射后蝗虫体内SOD、POD和CAT 活性以及MDA含量有明显变化。西藏飞蝗能够在光照逆境的条件下, 身体内的自由基清除系统和体内的抗氧化酶协调作用形成抗逆机制, 适应光照引起的环境变化。在测量不同光照下西藏飞蝗体内的过氧化物丙二醛(MDA)含量的实验, 说明蝗虫体内的过氧化反应与光照的时长、光的种类都有一定的联系。西藏飞蝗能够在青藏高原干燥的环境下生存, 是因为体内的抗氧化酶活性、含量和比例适应环境发生了改变。
4.3 西藏飞蝗受低氧的影响
西藏飞蝗的生存离不开氧气(O), 当O过低时会导致组织氧化损伤。为适应青藏高原含氧量低的环境, 西藏飞蝗的呼吸模式中存在不连续气体交换循环(DGC)呼吸模式。研究表明DGC呼吸模式与昆虫呼吸失水性和缺氧的环境中进化有关;西藏飞蝗通过增加DGC呼吸速率吸收更多的氧气(O)释放出二氧化碳(CO)。邓晓悦等通过测量和计算西藏飞蝗与东亚飞蝗的呼吸特征发现西藏飞蝗可以缩短DGC开放阶段, 用来补偿DGC频率增强带来的呼吸性水(HO)的损失。进一步证明西藏飞蝗的DGC呼吸模式与高原环境有关。
5 西藏飞蝗的综合防治措施
蝗灾被列为我国农业上的三大自然灾难之一, 严重影响我国的农牧业经济发展。在我国分布有三个种群的蝗虫, 其中西藏飞蝗主要分布在青藏高原地区, 为害牧草和禾本科农作物。我国对蝗虫的防治工作从以前的迷信、盲目治理到科学、安全、有效地防治, 到现在的生物技术防治、农业综合防治, 坚持植物资源保护、生物资源保护、环境资源保护以及生态资源保护的理念, 开发绿色可持续防治技术。
5.1 监测技术研究进展
西藏飞蝗的迁飞能力强, 种群规模大, 导致在监测上造成很大的问题。20世纪初, 我国通过鉴定蝗虫种类、研究蝗虫的生长发育以及调查蝗虫分布区域结合气象因子分析蝗虫发生规律从而建立检测站观测。随着我国计算机技术的快速发展, 对蝗虫的检测技术有很大的进步, 在20世纪末我国开始利用遥感(RS)检测蝗虫, 利用地理信息系统(GIS)预测蝗虫发生期和发生地点, 预防蝗灾。到21世纪, 我国开始将野外调查和3S技术相融合, 雷达技术、高空灯和地面诱捕相结合;利用大气环流建立我国蝗灾的检测预警体系, 实现蝗灾的提前预测并制定相对完善防治措施。
5.2 防治技术研究进展
针对西藏飞蝗的发生规律和为害症状结合生态安全建设, 防控技术从以前单一的物理防治或者化学防治应急防治策略发展到现在的以生物防治和农业防治多项防控技术相结合的绿色可持续防治体系。防治技术的不断改进与完善体现了人们想要控制西藏飞蝗的种群数量来调节西藏飞蝗的发生频率, 减轻西藏飞蝗对农业和畜牧业的影响。随着社会的进步, 人们在实践中挖掘更经济、环保的防治措施。
在我国传统防治蝗虫技术中一般人们通过饲养鸡、鸭或者其他蝗虫天敌来捕食蝗虫, 一直从古延续至今, 具有绿色、环保的优点, 但当大面积发生蝗虫时, 无法得到有效控制。经过人们多年的尝试, 现在我国对蝗虫的防治措施是采用常用的化学药剂对蝗虫进行防治, 这是蝗灾发生时首选的防治手段, 具有见效快、成本低、防治效果好等优点。由于单一追求防治效果严重引起了环境污染, 化学药剂使用方法技术不普及, 导致化学药剂在农户手中不能够正确使用, 造成水污染、大气污染、土壤板结等环境问题, 导致区域生态失衡, 物种多样性骤减。化学药剂的长期使用, 不但增加西藏飞蝗的抗药性, 而且化学药剂具有难降解的特性, 会经生物链和生物富集进入人体, 严重损害人体健康, 甚至有些化学药剂参与地球化学物质循环, 严重破坏地球生态系统。寻求一种绿色、环保、安全、高效的防治技术是目前绿色可持续防治蝗灾的首要任务。
21世纪以来, 我国对害虫的防治遵循不破坏生态环境和生物多样性的原则, 对传统的防控技术进行改进。在探索过程中对生态环境友好的生物防治技术登上历史舞台, 因为该防治方法对蝗虫不产生抗体, 且具有防治效果持久等特点。早在20世纪90年代, 蝗虫生物防治技术主要开发寄生性天敌的应用和植物源农药的开发利用两种主要形式, 通过人们不懈努力研发出一系列的杀蝗细菌、杀蝗真菌、杀蝗病毒、蝗微孢子虫、苏云杆菌等病原微生物菌剂, 多项杀蝗剂的研发在蝗虫的生物防治中起到十分重要的作用。随着研究的深入, 人们发现植物次生代谢物具有间接或直接的杀虫性, 有希望替代化学药剂防治蝗虫, 推动蝗虫生物防治, 将低耗、多元化、绿色的发展方向与生物制剂、化合物制剂、微生物制剂、天敌生物制剂等协同应用, 实现西藏飞蝗的绿色可持续控制。
在我国提出“预防为主, 综合防治”的植保方针以后, 西藏飞蝗得到有效控制, 生态环境破坏也得到缓解。随着“绿色可持续防控害虫”技术的提出, 西藏飞蝗的绿色防控技术也在不断开发。目前在明确西藏飞蝗发生规律与植被物候期的关系的前提下, 利用现代发达的监测预警技术研究飞蝗种群迁徙路线及规律, 从而建立短期、中期及长期的实时监测预警体系。保证生态安全的前提下, 建立集多项技术的绿色可持续防控技术体系。深度研究西藏飞蝗与寄主、非寄主、天敌之间的关系, 利用自然界存在的“推—拉策略”对西藏飞蝗防控符合我国目前科学的植保方针。
6 结语
西藏飞蝗能够适应全球气温变暖, 所以加强对西藏飞蝗实施绿色可持续防治技术依然是一项持续的课题。在青藏高原地区必须大力发展以生态治理为核心的环保控蝗技术, 利用RS、GIS、GPS等先进技术监控西藏飞蝗的迁徙。结合西藏自治区生态系统保护政策和当地的民俗建立应急防治策略和可持续治理策略相结合防控技术。利用现代科技研究产卵聚集信息素、聚集信息素和昆虫生长调节剂等干扰蝗虫的生命活动, 防止西藏飞蝗种群暴发。研究寄主植物中的化学物质对西藏飞蝗的趋势或非寄主植物中化学物质的避势, 为西藏飞蝗的绿色可持续防治探索新的方向。
猜你喜欢
北方音乐(2019年8期)2019-10-21
环球时报(2019-06-26)2019-06-26
故事作文·低年级(2019年6期)2019-06-20
时代英语·高一(2019年1期)2019-03-13
小猕猴智力画刊(2017年12期)2017-12-27
幼儿教育·父母孩子版(2017年10期)2017-12-13
现代农业科技(2017年13期)2017-08-09
农业科技与装备(2017年1期)2017-06-09
现代园艺(2016年7期)2017-01-09
现代园艺(2016年9期)2016-07-16
