
马产乳性状相关候选基因研究进展
2022-09-14 02:36李媛媛杨涛赵春江
中国畜牧杂志 2022年9期
李媛媛,杨涛,赵春江*
(1.中国农业大学动物科学技术学院,北京 100193;2.中国农业大学马研究中心,北京 100193)
马乳作为一种具有营养和治疗功效的饮品而受到人们关注。马乳饮品在我国、蒙古国、俄罗斯以及欧洲的一些地区广受欢迎。马乳中含有丰富的多不饱和脂肪酸和维生素,脂肪和胆固醇含量较低。马乳除具有较高的营养价值,还有美容、治疗疾病等功效。挖掘和验证马泌乳相关功能基因可为乳用马分子育种技术的开发和利用打下坚实的基础,对我国高产乳用马培育和马乳产业发展发挥重要的技术支撑作用,使我国早日育成泌乳性能优良的专门化乳用马品种。此外,随着生活水平的提高,人们越来越青睐健康、绿色的食品,有效开发和利用马乳资源,符合社会发展的需求,具有广阔的市场前景。本文介绍了马乳的营养特点和治疗功能,并着重对马泌乳性能、产乳性状相关候选基因和多组学技术在本领域研究中的应用进行了阐述。
1 马乳的特点和功能
马乳是由水、蛋白质、乳糖、乳脂、矿物质、酶等物质组成的白色或乳白色的胶状溶液。与奶牛、奶山羊等家畜相比,马乳中化学成分更接近人乳(表1)。马乳中干物质含量为11%,各种干物质成分如蛋白质、乳糖、乳脂、矿物质等的分散方式有所不同,分别以胶体形式、分子形式、浊液形式以及离子形式存在。
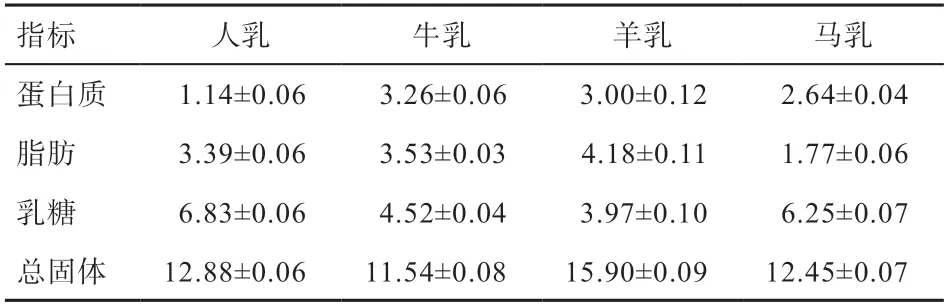
表1 牛、山羊、母马、母乳的主要成分含量[5] %
牛奶是目前最为普遍的乳品,但存在乳蛋白不易消化、部分人群对牛奶过敏等问题。与牛奶相比,马乳更容易被消化利用。马乳是乳清蛋白型乳,牛乳属于酪蛋白型乳,乳清蛋白型乳含有较多的可溶性蛋白,更容易被机体吸收。马乳中的酪蛋白遇酸会沉降为细小絮状,易溶于水,而牛奶中的酪蛋白遇酸会沉淀,几乎不溶于水。马乳中的乳糖属于易消化的糖类,在乳糖酶作用下可分解成葡萄糖和半乳糖,消化吸收率可达到98%以上。此外,马乳中的乳糖含量较高,是牛乳的1.5 倍以上。马乳中的脂肪含量虽然比牛乳低,但马乳中易消化吸收的不饱和脂肪酸和低分子脂肪酸含量是牛乳中的4~5 倍。马乳中含有丰富的甘油三酯酸,有利于人体健康。
除此之外,马乳还含有大量的生物活性蛋白,具有治疗疾病的食疗功效。长期有规律地饮用马乳能够缓解克罗恩病、肝炎、胃溃疡等炎症症状。马乳中含有婴幼儿生长发育所需要的全部营养物质,加之易消化吸收和低过敏性,因此适宜作为婴幼儿、老人和患有牛奶过敏症等特殊人群的乳品。随着马乳产品的不断研究和开发,马乳制品也在乳品市场崭露头角,受到欢迎。
2 马的泌乳性能
2.1 马的泌乳特点 母马每次泌乳大致分为2 个阶段:第1 个阶段为挤奶或者幼驹吮吸刺激下的泌乳初始阶段,母马将乳房中的乳液放完代表着这一阶段结束。马的蓄乳池较小,蓄乳能力差,第1 阶段的泌乳量约占总泌乳量的1/10,需15~20 s 乳汁即放完,然后乳汁停止分泌15~50 s,从而激活乳通路周围的平滑肌。第2 阶段开始于乳通路周围的平滑肌收缩,促使乳汁从乳导管进入乳池,迅速充满乳房,乳汁强烈分泌,每秒可分泌50~60 g,时间一般不超过1.5 min。此时挤乳动作要迅速灵活,若挤奶较慢,超过1.5 min,平滑肌会停止收缩,放奶停滞,造成大部分乳汁残留在乳房里。正常放乳情况下,母马可排出90%以上的乳汁。
由于母马是昼夜泌乳,泌乳量大,但乳房容积小,蓄乳能力差,只有在一定时间内多次挤,才能保证充分泌乳,因此需要多次挤奶。母马挤奶的间隔时间比较短,一般为2~2.5 h,每天可挤奶4~6 次。白天挤奶,夜里让马驹自由哺乳,挤乳和哺乳相结合,这是马乳生产的主要方式。另外,由于马泌乳的特点,母马放乳应尽量将乳房排空,否则会导致排乳抑制,影响泌乳量。
2.2 体型结构与泌乳性能的关系 体型外貌结构是影响马泌乳性状的重要指标之一。不同用途的马具有不同的体型外貌结构,但有关马体型结构与泌乳性能关系的研究较少,在奶牛中研究较多。奶牛的性能评估主要从体型外貌结构和产奶性能两个方面进行评定,其中体型外貌评价是奶牛育种中重要的选择指标。奶牛的体斜长、胸深、腰角宽、腹部大乳静脉弯曲程度等体型特征与产奶量有直接关系。马与牛都同属于哺乳类大家畜,体型外貌与泌乳性能之间的关系可能相似。
2.3 营养和环境对于马泌乳性能的影响 根据母马泌乳期特点进行科学饲养可提高泌乳量和乳品质。Doreau等研究发现,与饲喂95% 干草+5% 精料补充料的粗料型日粮相比,饲喂50% 干草+50% 精料补充料的精料型日粮组乳和血液中脂肪、蛋白和亚麻酸含量较高,但亚油酸含量较低。马乳产量与日粮中的精料量密切相关。母马的日常管理中要保证马厩卫生,确保母马有良好的饲养环境和一定的运动量,关注母马健康情况以保障马的泌乳量和乳品质。
3 马乳用性状候选基因研究进展
目前关于奶牛、奶山羊乳用性状相关基因的研究较多,而对于马乳用性状相关基因的研究相对较少,且多集中于研究马的表型数据,如马的泌乳规律和泌乳特点等。近年来,研究者对于马属动物的催乳素(Prolactin,)及催乳素受体(Prolactin Receptor,)、二酰甘油酰基转移酶(Diacylglycerol Acyltransferase,)、酪蛋白(Whole Casein)、生长激素受体(Growth hormone receptor,)、真核翻译延伸因子1D(Eukaryotic Translation Elongation Factor 1 Delta,)、甘油-3-磷酸酰基转移酶(Glycerol-3-Phosphate Acyltransferase Mitochondrial,)等基因的序列结构、基因多态性与马泌乳性状的相关性做了研究。
3.1 PRL 与PRLR PRL 是垂体前叶肽类激素,PRLR是与催乳素相结合,对乳蛋白基因表达起重要调控作用的蛋白。研究表明基因的遗传变异与产奶性能相关。张佳兰通过PCR-SSCP 技术和DNA 测序发现,奶牛外显子3 和7 上都有与产奶量和乳脂率显著相关的单核苷酸多态性位点(Single nucleotide polymprphism,SNP),其中外显子3 上AB 基因型的产奶量高,AA 基因型乳脂率高,外显子7 上BB 基因型具有更高的产奶量和乳脂率。Sirja 等通过PCRRFLP(Cleaved Amplification Polymorphism Sequence-Tagged Sites)和PCR-SSCP(Polymerase Chain Reaction-Single-Strand Conformation Polymorphism)方法在编码区和调控区挖掘到多个变异位点均与奶牛产奶性能相关,如奶牛PRL 基因外显子3 中的多态位点与乳脂肪和产奶量有关,可以作为与产乳性能相关的分子标记。Brym 等研究发现娟姗牛内含子9 的205 pb处存在一个A→C 的单碱基突变,CC 基因型娟姗牛的乳蛋白含量较高。此外Sirja 等在苏格兰奶牛中发现了蛋白前体第18 处丝氨基酸变为天冬酰胺的突变,该突变会影响奶牛乳脂率、蛋白量和产奶量,特别是对乳脂率和蛋白量的影响很大。在奶山羊研究中发现基因外显子存在SNP,其中一些突变与山羊的产奶量相关。姚新奎等在马的研究中发现,该基因第4、5 外显子存在多态性。此外,高程程等对伊犁马基因进行多态性检测,发现该基因侧翼区存在多态性,均与产奶量有关。根据基因的研究进展,可将其作为研究马泌乳性能的候选基因。
3.2 DGAT家族在机体沉积脂肪的部位均有表达,研究表明其与家畜产乳性状有关。根据细胞定位和结构差异,将分为和蜡酯合成/ 二酰甘油酰基转移酶()。是甘油三脂合成过程中最后一步反应的限制酶,在牛中该基因位于的着丝粒末端,在泌乳性状相关的数量性状基因位点(Quantitative Trait Locus,)定位区间内。已有研究表明,奶牛基因第8 外显子中存在一个双碱基突变,此突变使第232 位的赖氨酸变为丙氨酸(K232A),从而影响乳脂率。徐秀荣等研究发现奶牛基因K232A 突变型个体的乳脂率比野生型高。Spelman 等在新西兰3个奶牛品种的K232A 位点的研究中发现等位基因K 和高乳脂率相关。李聪研究表明,基因第8 外显子中存在AA→GC 的双核苷酸突变,该错义突变造成所编码的赖氨酸变为丙氨酸,并对奶牛的泌乳性能尤其是乳脂率造成影响。在马属动物驴的研究中,肖海霞等、王颜颜使用RACE 技术得到基因3'端的部分序列,还对基因第5,6 外显子和第3,4,5 内含子的多态性进行检测,发现一些与驴体高、胸围、体长、净肉率等性状相关的突变,而基因对马产乳性状相关性的研究尚未见报道。
3.3 酪蛋白基因 酪蛋白是由乳腺上皮细胞合成的一组蛋白,是乳蛋白的主要组成部分,主要包括-酪蛋白(Casein Alpha S1,)、-酪蛋白(Casein Alpha S2,)、-酪蛋白(Casein Beta,)和-酪蛋白(Casein Kappa,)4 种类型。在过去的几十年里,对反刍动物以外家畜的酪蛋白基因研究已积累了一定成果。最近在美洲驼、骆驼和家马中发现了许多酪蛋白基因多态性位点。Jakub 等在马的研究中筛选了酪蛋白基因5' 侧翼区多态性,共检测到23 个多态性位点,其中包括21 个单核苷酸多态性()和2 个新的插入缺失标记(Indels),超过1/3的位点是在基因序列中发现的。表2 展示了马酪蛋白基因5'-侧翼区多态性检测的研究结果,大部分基因变异在各马种中分布广泛。在基因的相应区域中发现了3 个基因变异。基因的1个突变(c.-1732A>G 多态性)仅在胡克尔马(HUC)和西莱西亚马(SIL)2 个品种中检测到。进一步分析显示,检测到的22 个多态性位点中有17 个可以改变转录因子预测的结合位点,这些突变点可能对基因表达和乳成分产生影响。此外在马属动物的研究中还发现,基因多态性与产奶性能有着密切关系。Hobor 等研究发现马基因的第1 和第4 外显子区各有2 个。但在对疆岳驴基因的研究中没有在第1外显子区域发现多态性,这可能是由于马、驴种属差异造成的。

表2 马酪蛋白基因5'-侧翼区多态性[28]
在奶山羊研究中,乐祥鹏对奶山羊基因外显子2、9、12、17 和19 与内含子14 进行了多态性检测,发现基因第9 外显子的多态性与产奶量相关;基因第14 内含子也有一个与产奶量相关的突变。蓝贤勇等发现西农萨能奶山羊基因的片段中存在突变,不同基因型对平均产奶量有显著影响。此外,在付小波等在荷斯坦牛对基因的研究中发现,荷斯坦奶牛和黄牛改良品种的基因5' 端序列的突变与产奶量有关;在基因第2 外显子检测到一个突变,形成AA、AB 和BB 等3 种基因型,其中AA 型乳蛋白率较高,同时黄牛改良品种基因第2 外显子的AA 基因型个体的乳脂率和乳蛋白率显著高于BB 和AB 基因型个体,AA 基因型个体产奶量也高于BB 和AB 基因型个体。
由此可见,酪蛋白基因多态性与马、奶山羊、奶牛等家畜的泌乳性能都具有一定相关性,因此酪蛋白基因可作为影响泌乳性状的候选基因。
3.4是生长激素(Growth Hormone,)的受体蛋白,是细胞因子和造血因子受体超家族的成员之一。基因的碱基序列发生改变会影响的正常发挥,从而影响奶牛产奶、产肉等性状。Rahmatalla等在荷斯坦牛中发现基因的第836 位T →A突变与产奶量相关。王凯等、邱峥艳等研究表明荷斯坦奶牛基因多态性对乳脂率有显著影响,但对产奶量影响不大。马彦男等验证了奶牛基因F270Y 突变位点的功能,证实了AA 型奶牛产奶量高于AT 型。马妍等对荷斯坦奶牛基因与泌乳性能相关的研究表明,奶牛基因F279Y 位点TT 与AT 基因型个体在产奶量和乳脂率方面差异显著。但也有一些研究发现基因多态性与产奶量并没有显著相关性。马的生长激素受体基因位于第21 号染色体上,有10 个外显子,但目前关于马基因与泌乳性状相关性研究的报道较少。孟拮等对伊犁马基因的多态性进行检测,在启动子区发现了1 个SNP 位点,该突变对产奶量有一定的影响,并认为该位点有望应用于乳用马的选育。
3.5 EEF1D是翻译延伸因子复合物的的亚基。谢岩发现牛基因有4 种剪切体,对该基因进行功能验证发现基因上存在一个与乳脂率显著相关的多态位点。Zhang 等在奶山羊的研究中发现具有两种可变剪切体,其研究结果同样也表明该基因与乳脂合成有关。刘轩等在荷斯坦奶牛的研究中通过关联性分析发现基因启动子和UTR 区上的SNPs 与产奶量、乳脂率和乳蛋白率相关。蒋秋斐等发现牛基因的5' 非翻译区存在一个突变位点,该位点能够影响乳脂的分泌。通过进行转录因子预测并结合启动子活性分析,发现该突变会引起基因的表达上调,最终导致转录因子Sp3转录起始位点后移33 bp。突变型的活性大约是野生型的3 倍,但Sp3 转录起始位点变化对乳脂含量的调控机制还需深入的研究。目前关于基因与乳脂率相关的研究在牛、羊等家畜中开展较多,但该基因在马属动物上尚缺乏研究。
3.6 GPAT 甘油-3-磷酸酰基转移酶()脂肪族酰基为底物,催化甘油-3-磷酸脂酰基化反应,产物为溶血磷脂酸。酶活性较低,是脂质合成代谢途径中的限速酶。有两个同工酶亚群,分别是微粒体和线粒体。其中线粒体又简称为。是合成甘油三酯的第一步反应的关键酶,能够影响乳脂的形成过程。赵尧璐研究发 现基因3 个SNP位点E20-3386G>A、I18-652A>G 和I18-726A>G 均与奶牛产奶性状有关,其中E20-3386G>A 位点与奶牛乳蛋白率及体细胞数有显著相关;I18-652A>G 位点与奶牛乳蛋白、干物质和体细胞数相关;I18-726A>G 位点与奶牛蛋白率、产奶量、干物质、体细胞数均显著相关。多个研究均表明基因与奶牛乳脂的合成有关。此外,基因还能够影响肉牛的肉质性状,研究表明该基因对脂质代谢的调控起到重要作用,并且对改善肉质有明显的效果。除了在牛上的研究,目前还未有基因与马及其他家畜泌乳性能相关性的报道。
除了上述基因外,等基因也对家畜的泌乳性能有影响,但这些基因在马属动物中的研究都比较少,大多数研究集中在奶牛、奶山羊等传统泌乳家畜上。在今后的研究中关于马泌乳性状相关候选基因的挖掘还有很多工作尚待完成。
4 多组学技术在乳用性状基因研究中的应用
近十几年来,随着组学技术的迅猛发展,利用多层次组学的信息,从基因组、转录组和蛋白组水平挖掘重要性状的功能基因,已成为家畜遗传学研究的有力手段。通过组学方法研究马的泌乳性能调控基因及其遗传机制是将来重要的研究方向。这方面马属家畜研究成果尚少,但可以以奶牛的相关研究作为参照。
目前全基因组关联分析(Gene Association Study,GWAS)已广泛应用于奶牛产乳性状候选基因的筛选中。如Li采用单标记传递不平衡检验(LI-TDT)和单标记回归分析(MMRA)2 种统计分析方法对荷斯坦奶牛产奶性状进行GWAS 分析,发现105 个SNPs 达到全基因组显著水平,其中38 个SNPs 被2 种方法同时检测到,其中和两个SNPs 分别位于影响产奶量和乳脂率的明星功能基因DGAT1 和GHR 附近,由此判断奶牛产奶量和乳脂率可能受这2 个SNPs 的影响。Nayeri 等对加拿大荷斯坦牛乳成分性状开展GWAS 研究,鉴定出与产奶性状相关的候选基因和通路,包括等。Nayeri 等使用777K SNP 芯片进行单标记回归全基因组关联分析,鉴定出与产奶性状显著关联的基因有和。Chen 等对1 092 头荷斯坦牛进行简化基因组测序,GWAS 结果显示17 个单倍型与产奶性状显著关联,最后筛选到和等基因与产奶量显著关联。Li 等采用Illumina 牛54K 芯片对784 头中国荷斯坦奶牛中的22 种乳脂肪酸进行了全基因组关联研究,共检测到83 个全基因组显著SNPs 和314 个与乳脂肪酸性状相关的显著SNPs。
在转录组学研究方面,乳腺组织和乳腺上皮细胞等不同来源的RNA 已广泛应用于转录组测序研究,并鉴定出与其相关的新转录本和非编码RNA。Yang 等针对干奶期和泌乳初期的血液以及泌乳初期和高峰期的牛奶样品,分析了6 头高低产的中国荷斯坦奶牛的转录组样本,共鉴定出1 408 和1 459 个差异表达的基因,其中有363 个差异表达基因位于已知的产奶性状QTL 区域。针对奶牛不同的泌乳期,Wickramasinghe 等开展了泌乳初期(D15)、高峰期(D90)和末期(D250)3 个时期的乳腺细胞转录组测序分析,分别挖掘出16 892、19 094 和18 070 个差异表达基因,富集分析发现泌乳初期和高峰期的高表达基因主要集中在脂类乳合成通路中,而编码酪蛋白、乳清蛋白和乳糖酶基因表达则主要集中在泌乳初期。
在蛋白组学研究方面,Mol 等对印度牛3 个不同的泌乳期(初期、中期和末期)牛奶的蛋白质组特征进行比较分析,鉴定出403 个差异表达的蛋白,主要富集到乳合成通路中。Janjanam 等用双向电泳技术针对3 个泌乳期(初期、高峰期和末期)的高产奶和低产奶的8 头奶牛奶中乳腺上皮细胞进行蛋白组比较分析,发现一些影响产奶量和泌乳持久力的关键蛋白,在产奶量高低组共挖掘到22 个差异蛋白,且注释分析显示大部分差异蛋白与乳合成通路、催化和结合活性等相关。
由上述可见,组学分析技术是乳用性状相关基因挖掘的有力研究手段,对探究家畜乳用性状发生的遗传基础具有重要意义。目前,尚缺乏与马产乳性状相关的基因组、转录组和蛋白组分析的研究,但在奶牛等乳用家畜上的研究结果表明,充分利用多组学技术的优势,解析与目标性状相关的基因和因果突变是一种有效的研究策略。基于多组学数据的研究策略可以大幅度提高候选基因筛选的可靠性,提高研究效率,从而揭示马产奶性状的分子遗传机制。同时,由于奶牛中相关研究已积累了大量成果,可通过比较基因组学的方法,在马产奶性状相关基因组学分析中充分借鉴和利用奶牛中已有的研究成果。
5 总结
马乳由于其特殊的理化性质而受到人们的欢迎,马乳生产和加工具有广阔的市场前景。尽管马属动物泌乳相关基因尚缺乏系统研究,但通过对奶牛及其他家畜的长期遗传学研究,人们已经筛选并鉴定了许多与泌乳性能相关的重要功能基因,这些研究成果为马属动物的同类研究提供了重要研究基础和参考依据。
猜你喜欢
中国典型病例大全(2022年9期)2022-04-19
现代畜牧科技(2018年5期)2018-05-14
现代畜牧科技(2018年5期)2018-05-14
医学信息(2017年1期)2017-02-28
科技与创新(2015年21期)2015-12-01
现代畜牧科技(2015年12期)2015-10-21
湖北畜牧兽医(2015年3期)2015-06-05
湖北畜牧兽医(2014年9期)2014-12-02
中国民族民间医药·下半月(2014年5期)2014-12-02
湖南饲料(2014年4期)2014-08-26
