
青蒿琥酯对人脐带间充质干细胞诱导的去分化肌成纤维细胞凋亡的影响
2022-09-13 08:53王雅文吴苗耿素霞曾令基王玉连赖沛龙杜欣翁建宇
实用医学杂志 2022年13期
王雅文 吴苗 耿素霞 曾令基 王玉连 赖沛龙 杜欣 翁建宇
1华南理工大学医学院(广州 510006);2广东省人民医院(广东省医学科学院)(广州 510080)
闭塞性细支气管炎(bronchiolitis obliterans,BO)是异基因造血干细胞移植后的肺部严重并发症,小气道纤维疤痕形成是BO 不可逆的病理损伤,5年死亡率高达50%,严重影响患者的生活质量及寿命。现有抗炎、抑制受体酪氨酸激酶的抗纤维化治疗方式仅能轻微提高纤维化患者的生活质量和生存率,因此探索新的治疗策略极为迫切[1-3]。肌成纤维细胞(myofibroblast,MFB)是纤维化的“核心效应细胞”,是细胞外基质的“生产车间”[4]。MFB 分化/转化失调、去分化紊乱、增殖失控和凋亡逃逸是纤维化发生发展的重要机制[5-7]。MFB 的来源多样性、异质性和调控机制复杂性导致迄今还未开发出强有力的抗纤维化药物,因此对MFB 调控的研究将成为治疗纤维化的新方向。间充质干细胞(mesenchymal stem cells,MSC)抗纤维化作用被越来越被多地探索和报道,大量临床前研究显示MSC 或MSC 来源囊泡具有抗纤维化作用[8-10]。MSC 通过免疫调节、抗炎和抑制FB 分化、上皮细胞/内皮细胞转分化等机制在纤维化的多个环节发挥抗纤维化作用,但MSC 对MFB 的具体作用仍不清楚。MSC 在抗纤维化的临床应用研究进展缓慢,疗效仅部分延缓进展。如何提高MSC 抗纤维化作用一直是研究领域的热点。
青蒿琥酯(artesunate,ART)可诱导MFB 凋亡,从而发挥抗纤维化的作用[11-12]。本研究团队前期工作也证实了青蒿琥酯可通过调节BCL2 家族蛋白诱导肺MFB 凋亡[13]。但青蒿琥酯长期使用具有肝毒性、肾毒性等副作用,使其在疟疾以外疾病的应用受限制[14-15]。青蒿琥酯联合MSC 是否可以提高抗纤维化作用,克服MSC 疗效不稳定、减少青蒿琥酯不良反应的不足尚未明确。
本研究将观察人脐带间充质干细胞(hUCMSC)对肺MFB 的具体作用,探索青蒿琥酯联合hUCMSC 对MFB 增殖、凋亡、细胞外基质合成功能的影响,为青蒿琥酯联合MSC治疗BO提供实验依据。
1 材料与方法
1.1 材料α-MEM 培养基、胎牛血清、胰酶(Gibco公司),TGFβ1(PeroTech 公司),青蒿琥酯(Sigma公司),CCK8 试剂盒、凋亡试剂盒(广州松鼠生物),逆转录及PCR 试剂盒(湖南艾科瑞生物),流式抗体(BD),人胚肺成纤维细胞(MRC-5)(美国ATCC 公司),hUCMSC 培养基及诱导分化试剂盒(赛业)。
1.2 方法
1.2.1 hUC-MSC 制备足月产妇签署脐带捐赠知情同意书后,无菌取人脐带样本10 cm。分离华通氏胶,剪成1~2 mm3碎块,加入Ⅱ型胶原酶,消化6 h。于37 ℃培养,每3 天换液,当细胞融合达80%~90%,按1∶3传代。实验所用细胞均为P3 代。
1.2.2 hUC-MSC 鉴定(1)表面标志物检测:于流式管中加CD90/HLADR-FITC、CD73/CD34-PE、CD105/CD45-APC 各5 μL,室温避光孵育30 min,流式上机操作。(2)分化能力检测:以5 × 104/孔将MSCs 铺至6 孔板,当细胞融合度达70%时,更换为成骨诱导培养基,每3 天换液1 次;当细胞融合度达100%时,更换为成脂诱导培养基。约21 d 后,进行油红O 和茜素红染色。
1.2.3 肌成纤维细胞制备P4-P7 代MRC-5 以2×104/孔接种于12孔板,各孔中加入10 ng/mL TGF-β1诱导48 h,检测α-SMA 表达水平。
1.2.4 实验分组(1)FB 组(MRC-5):FB、FB-丙酮组、ART-FB 组、MSC-FB 组、ART-MSC-FB 组;(2)MFB 组(MRC-5+TGF-β1):MFB 组、MFB-丙酮组、ART-MFB 组、MSC-MFB 组、ART-MSC-MFB 组;(3)MSC-deMFB 组:MSC-deMFB 组、MSC-deMFBTGFβ1 组、MSC-deMFB-MSC 组、MSC-deMFB-TGFβ 1-MSC。MRC-5 以2 × 104/孔接种于12 孔板,当FB融合达60%左右时,加入TGF-β1诱导48 h后,分别加入100 μmol/L ART、MSC或ART+MSC各24 h。在12 孔板上方放置0.4 μm 的小室,实验组的小室接种2 × 103/孔的MSC,500 μL/孔,对照组的小室添加相同体积的培养基。
1.2.5 细胞增殖检测hUC-MSCs 与MFB transwell共培养24 h,PBS洗涤,加10%CCK8培养基,于37 ℃培养箱中避光孵育1 h,测定450 nm处的OD值。
1.2.6 细胞凋亡的流式检测按照1.2.4 实验分组进行分组,于流式管中加入Annexin-V 和PI 各5 μL,避光孵育5 min,流式上机。
1.2.7 qPCR检测基因mRNA表达采用Trizol 法提取细胞总RNA,按说明书合成cDNA及PCR体系,引物序列见表1。数据以2-ΔΔCt计算α-SMA、COL1A1及FN1 的相对表达量。

表1 PCR 引物序列Tab.1 Primer sequences for PCR
1.2.8 WB 检测基因蛋白表达采用RIPA 法提取细胞总蛋白,进行变性。蛋白上样量为每孔30 μg,电泳90 min,电转2 h,封闭1 h,TBST 洗三次。一抗4 ℃孵育过夜,二抗室温孵育1 h,上机曝光,检测α-SMA、COL1A1 及FN1 的相对表达量。
1.3 统计学方法使用GraphPad Prism 9.0 对所有数据进行统计学处理,数据用均值±标准差表示,两组间比较采用独立样本t检验,多组间比较采用单因素方差分析。P<0.05 为差异有统计学意义。
2 结果
2.1 hUCMSCs的鉴定P3代hUCMSCs呈长梭形、多角形(图1A),可诱导成脂细胞(图1B)和成骨细胞(图1C)。hUCMSCs 表型符合国际细胞协会MSC标准(图1D)。
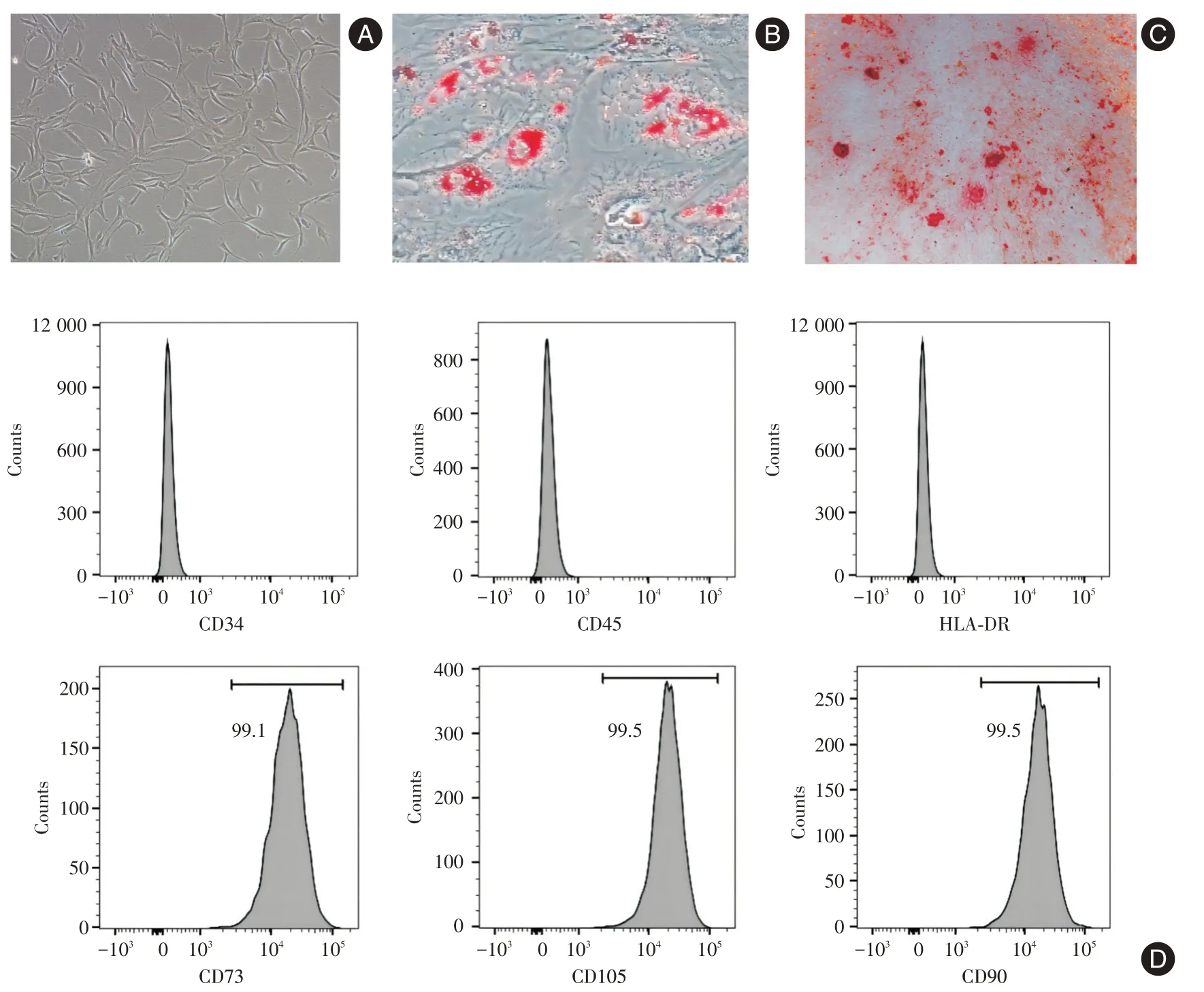
图1 hUCMSCs 的鉴定Fig.1 Identification of hUCMSCs
2.2 hUCMSCs 促进MFB 增殖当hUCMSCs 与MFB transwell 共培养24 h 后,MSC-MFB组细胞形态与MFB、FB 无明显差异,细胞生长更密集、呈紧密连接,形似网格状(图2A);CCK8 结果表明MSCMFB 组较MFB组,OD值增高(P<0.01,图2B)。
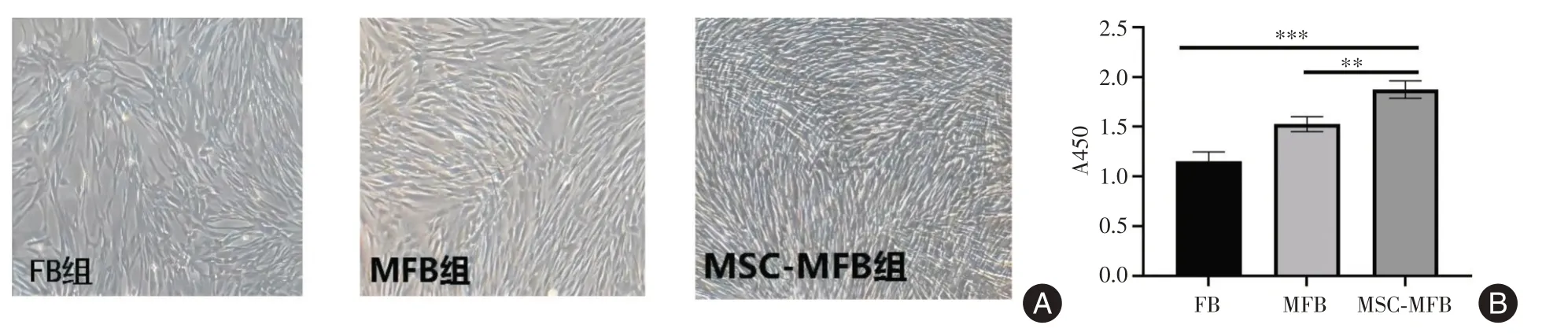
图2 MSC 促进肺MFB 增殖Fig.2 MSCs promote proliferation of myofibroblasts
2.3 hUCMSCs 诱导MFB 可逆性去分化当hUCMSCs 与MFB transwell 共培养24 h 后,α-SMA、COL1A1、FN1 的mRNA(图3A)和蛋白(图3B)表达水平均较MFB 组明显降低,提示hUCMSCs 可诱导MFB 去分化(MSC-deMFB)。
为了解hUCMSCs 对MFB 的去分化作用是否可逆,撤除MSC 后,MSC-deMFB 经TGF-β1 重新诱导后,α-SMA 及细胞外基质的mRNA(图3C)和蛋白(图3D)表达水平均增加,并恢复至MFB 水平。上述结果提示MSC诱导MFB的去分化作用是可逆的。

图3 MSCs 诱导MFB 可逆性去分化Fig.3 MSCs induce reversible dedifferentiation of myofibroblasts
2.4 ART 促进MSC-deMFB 凋亡CCK8 结果 显示:ART-MSC-MFB 组的细胞活性较ART-MFB 组降低(P<0.05,图4)。细胞的总凋亡率见表2。实验结果提示:(1)FB 对ART 诱导凋亡作用优于MFB(P<0.01);(2)ART 对MSC 诱导的去分化MFB(MSC-deMFB)的凋亡作用优于ART 单独作用于MFB(P<0.05)。

图4 ART 促进de-MFB 凋亡Fig.4 ART promotes de-MFB apoptosis
表2 细胞总凋亡率Tab.2 Total apoptosis rate±s

表2 细胞总凋亡率Tab.2 Total apoptosis rate±s
组别FB 组ART-FB 组MFB 组MSC-MFB 组ART-MFB 组ART-MSC-MFB 组总凋亡率(%)0.67±0.06 11.23±0.89 0.90±0.07 0.88±0.03 5.98±0.15 8.45±0.27
2.5 ART 协同hUCMSCs 抑制MFB 的细胞外基质合成MSC、ART 均抑制MFB 的α-SMA、COL1A1的mRNA和蛋白表达;当ART联合MSC作用时,MFB的α-SMA、COL1A1表达水平进一步降低(P<0.01)。实验结果表明:ART 协同hUCMSCs 进一步抑制MFB 合成细胞外基质。见图5。

图5 ART 协同hUCMSCs 抑制MFB 合成细胞外基质Fig.5 ART synergizes with MSCs to inhibit extracellular matrix of the myofibroblasts
3 讨论
BO 的病因复杂、多环节失衡(气道上皮损伤→炎症/免疫介导→多种细胞向肌成纤维细胞转化→MFB 去分化失调、增殖失控、凋亡逃逸→纤维瘢痕形成)及周而复始的病理生理过程,最终发展为不可逆的细支气管狭窄、闭塞[16-18]。既往临床实践围绕抗炎、抑制纤维化信号通路活化(尼达尼布、吡非尼酮)的治疗方法,治疗效果不尽如人意。究其原因,传统治疗策略并不能切断复杂病因、不能阻断来源丰富的各种细胞向MFB 转化、不能抑制MFB 增殖和逆转MFB 凋亡逃逸,而且更无法消退已形成的气道疤痕[19-20]。
MSC 抗纤维化作用越来越被关注,既往研究重点集中在成纤维细胞、上皮细胞、内皮细胞等各种细胞向MFB 转化这一过程的调控[8,21-22],而对于纤维化核心细胞——MFB 的关注较少。
本研究发现当MSC 与MFB 共培养后,细胞呈现增殖状态,其纤维化相关基因的表达明显下调,恢复至接近FB 的起始水平,这种现象称之为“MFB 去分化”。本研究把经MSC 诱导的去分化MFB 称之为“MSC induced dedifferentiation of MFB,MSC-deMFB”(实验中的MSC-MFB组)以区分MFB和FB。为了进一步了解MSC 对MFB 的去分化作用是否可逆,本研究对MSC-deMFB 进一步分组实验,结果发现:当撤除MSC 的刺激后,加TGF-β1 再刺激,MSC-deMFB 可恢复MFB 的特征;而在MSC 持续作用下,MSC-deMFB 仍然保持去分化特征,尽管再加入TGF-β1 也不能逆转MSC 对MSC-deMFB 的持续去分化作用。上述结果解释了MSC 临床疗效不稳定、易复发的部分原因:MSC 诱导MFB 去分化作用是可逆的,而且依赖于MSC 的持续作用;临床纤维化的病因往往不易去除,如在TGF-β1/SMAD 信号通路持续活化的环境下,一旦撤除MSC,增殖的MSC-deMFB 向MFB 逆转(re-deMFB)势必导致更为剧烈的纤维化效应。本研究的发现为MSC 治疗纤维化疾病的临床研究设计提供理论依据。克服MSC 对MFB 增殖和可逆性去分化作用,将是提高其抗纤维化的重要策略。
近年来,研究证实了青蒿琥酯在肺、肾、肝等器官纤维化疾病中的抗纤维化作用[23-25],其作用机制为:(1)抗炎和免疫调节[26]:青蒿琥酯可抑制炎症细胞的活化及招募,抑制TGF-β、TNF-α 等促炎因子的表达;(2)促进凋亡:青蒿琥酯可通过细胞内在性途径与外在性途径促进细胞凋亡。本研究前期研究表明青蒿琥酯可通过调节BCL-2 家族蛋白的表达进而促进MFB 凋亡[13],从而发挥减轻cGVHD 的作用[27]。青蒿琥酯因抗疟作用被广泛使用,但即使在安全剂量范围内,也会引起血液系统疾病、听力障碍、肝脏毒性和心律不齐等不良反应[14,28]。青蒿琥酯联合MSC 治疗纤维化疾病是否能达到协同作用的目的:即是否可提高抗纤维化的疗效,又防止MSC 疗效反弹、减少青蒿琥酯的毒副作用。由此,笔者进一步探索了青蒿琥酯对MSC-deMFB 的作用。青蒿琥酯诱导凋亡作用强度依序如下:FB>MSC-deMFB>MFB;并可克服MSC对MFB 的增殖作用,增强MSC 抑制MFB 细胞外基质合成功能。由此笔者认为:青蒿琥酯可克服MSC诱导的MFB 增殖和可逆性去分化的不利现象,青蒿琥酯协同MSC 可能是有效安全的抗纤维化治疗新策略。
本研究首次揭示MSC 可促进MFB 增殖、诱导MFB 可逆性去分化,并首次证实青蒿琥酯可诱导MSC-deMFB 凋亡、协同MSC 抑制MFB 的细胞外基质合成。为BO 乃至纤维化疾病的治疗提供了新的理论和实验基础。但本研究也存在一定局限性:因体内环境和体外培养条件不同,须进行体内和临床研究加以验证;虽有研究认为MSC-EV 和MSC-SF 均有抑制FB 分化和ECM 合成作用,但MSC-EV 的作用更为稳定和持久,MSC-EV 中所含的miR21、miR29 参与抗纤维化调控[29];MSC 分泌物调控MFB 去分化的机制尚不清楚,有待进一步实验研究加以阐明。
猜你喜欢
现代建筑电气(2022年6期)2022-12-16
检验医学(2022年3期)2022-05-05
中国中医药信息杂志(2022年1期)2022-01-13
计算机时代(2021年9期)2021-10-08
东坡赤壁诗词(2021年4期)2021-09-05
保健医苑(2021年7期)2021-08-13
感染、炎症、修复(2021年1期)2021-07-28
世界科学技术-中医药现代化(2020年2期)2020-07-25
新世纪智能(高一语文)(2020年5期)2020-07-24
商品与质量(2019年27期)2019-12-02
