
大庆库里泡浮游植物群落时空变化及其与环境因子的相关性
2022-09-02 12:25朱以敏徐洪伟张殊闻李铭震余至立陆欣鑫范亚文
中国环境监测 2022年4期
朱以敏,徐洪伟,张殊闻,梁 铖,李铭震,余至立,熊 普,陆欣鑫,刘 妍,范亚文
1.黑龙江省水生生物多样性研究重点实验室,黑龙江 哈尔滨 150025 2.哈尔滨师范大学生命科学与技术学院,黑龙江 哈尔滨 150025
浮游植物作为水生态系统中食物链的开端及主要的初级生产力,同时也是水生态系统中物质循环、能量流动和信息传递的生态载体,其物种多样性的变化会直接影响到水生态系统的结构与功能[1]。浮游植物时空分布格局具有较显著的周期性规律,能表征出不同水体的生态状况和特点,因此,探究浮游植物群落的时空更替规律与氮磷营养盐含量的耦合关系,可为水体营养状态监测和水质管理提供理论支撑[2-3]。有研究表明,浮游植物群落结构的变化往往可作为分析湖库空间多样性变化模式的有效生态学信号[4]。受水体流速、径流量等水文特征及氮磷营养盐时空分布的影响,浮游植物的群落结构在湖泊的不同水文分区具有一定的差异[5-6]。
库里泡地处黑龙江省大庆市肇州县,位于松花江北岸,东西最宽处约5 km,南北最长处约16 km,湖体面积约81 km2,最大蓄水量约9.1×107m3,是串联在安肇新河上的最大排污型湖库[7]。大庆市人口众多且重工业较发达,区域内大部分生活污水和工业废水都经由安肇新河注入库里泡后排入松花江,因此,库里泡在对流域内营养盐的拦截方面起着重要作用。库里泡距离位于松花江下游的哈尔滨市四方台饮用水水源地仅184 km,其水体质量与哈尔滨市居民的生活用水安全息息相关[8]。然而近年来,随着大庆市工业规模的扩大,库里泡湖区承受的排污量增加明显,水体营养负荷加大[7]。根据2015—2018年《黑龙江省生态环境状况公报》提供的数据,库里泡水质类别连续4年均为劣V类,水体污染严重。库里泡作为典型的温带中小型湖库,相比于大型湖库,其纳污、缓冲和自净能力较弱[9]。
目前,浮游植物群落结构的动态变化已被广泛应用于国内外湖泊水体水环境质量评价[5-6,9],国内相关报道主要见于针对太湖[10]、洱海[11]、抚仙湖[12]、鄱阳湖[13]、查干湖[14]、乌梁素海[15]等湖泊开展的浮游植物群落结构分布和水质评价研究。近年来对库里泡浮游植物进行的一系列生态调查研究显示,库里泡浮游植物群落演替具有一定的季节特征。曹文钟等[7]于2012年5—10月连续对库里泡浮游植物进行了6次监测,发现其群落结构呈现为绿藻-硅藻型,并据此推断库里泡水体营养程度相对较高;杨双[16]在2014—2015年对库里泡浮游植物功能类群进行的研究中发现,库里泡春季水体质量较佳,而夏、秋两季水体均呈现出较高的富营养状况。目前,关于库里泡浮游植物群落结构时空变化的研究主要关注浮游植物的群落结构特征,对浮游植物群落时空分布格局与水体环境特征的相关性的研究尚处于本底数据挖掘和积累阶段。通过对库里泡浮游植物群落结构演替与水体环境因子状况的耦合关系进行研究,可以及时掌握水体营养状况及污染特征,为温带中小型湖泊生态修复、生境稳态保护及动态变化机制研究提供可靠的生态学及生物学基础数据和理论依据。
1 材料与方法
1.1 采样点设置
在库里泡设置了13个采样点(图1),其中,1#、2#采样点分布于入水口,3#、4#、5#、6#、7#、8#、9#、10#、11#采样点分布于湖泊区,12#、13#采样点分布于出水口。分别于2019年5月(春季)、8月(夏季)、10月(秋季)对13个采样点进行了样品采集。因冬季湖面结冰严重,未对该季节样品进行采集。
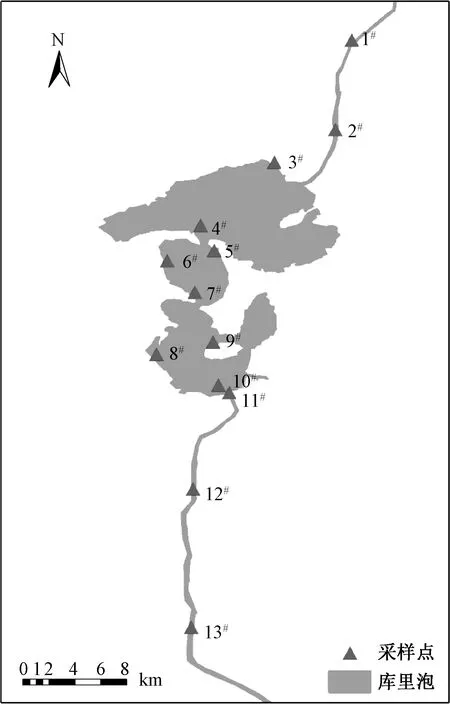
图1 库里泡采样点示意图Fig.1 Sample sites in Kulipao Lake
1.2 浮游植物采集、处理及鉴定
浮游植物定性样品采集方法:采用25#浮游网在水面下0.5 m左右处做“∞”形循环,将捞取到的浮游植物定性样品收集于采样瓶中,然后立即用4%的甲醛溶液进行固定。带回实验室后,使用光学显微镜进行种类鉴定。浮游植物定量样品采集方法:在水面下0.5 m左右处取1 L水样,用鲁格试剂对其进行固定、沉淀,最终浓缩至50 mL。在进行浮游植物鉴定和计数时,取0.1 mL样品注入浮游植物计数框,参考浮游植物鉴定相关文献[17],在10×40倍光学显微镜下进行物种鉴定和计数。
1.3 数据采集、处理与分析
1.3.1 理化指标的采集
使用多参数便携式水质分析仪(美国Visai,YSI Pro Plus)进行电导率、pH、溶解氧(DO)和水温数据的采集;于24 h内对水样总磷(TP)、总氮(TN)、高锰酸盐指数(CODMn)及五日生化需氧量(BOD5)进行实验室测定,测定方法参考《地表水环境质量标准》(GB 3838—2002)[18]。
1.3.2 优势种的判定
依据各采样点浮游植物优势度来判定优势种,计算公式如下:
(1)
式中:Y为优势度;Ni为第i个物种的数量;N为所有物种的总数量;fi为第i个物种在各采样点出现的频率。将Y值大于0.02的种类判定为优势种。
1.3.3 营养状态指数
采用营养状态指数对库里泡水体富营养状态进行评价[10],计算公式如下:
TLI(TN)=10×[5.453+1.552×
lnc(TN)/ln2.5]
(2)
TLI(TP)=10×[9.436+1.488×
lnc(TP)/ln2.5]
(3)
TLI(CODMn)=10×[0.109+2.438×
lnc(CODMn)/ln2.5]
(4)
TLI=1/3×[TLI(TN)+TLI(TP)+
TLI(CODMn)]
(5)
式中:TLI为营养状态指数;TLI(TN)、TLI(TP)、TLI(CODMn)为各污染物对应的营养状态分指数;c(TN)、c(TP)、c(CODMn)为各污染物的浓度。评价标准:0
1.3.4 物种多样性指数
对于浮游植物群落多样性,采用Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数和Simpson生态优势度指数进行评价[19],计算公式如下:
(6)
H=(S-1)/lnN
(7)
J=H′/lnS
(8)
(9)
式中:H′为Shannon-Wiener多样性指数;H为Margalef丰富度指数;J为Pielou均匀度指数;D为Simpson生态优势度指数;S为浮游植物物种数;Ni为第i种的数量;N为所有物种的总数量。
1.3.5 数据分析
本实验采样点地图使用Google Earth和ArcGIS 10.2软件绘制,浮游植物优势种与水环境变量间的冗余分析(RDA)使用CANOCO 4.5完成。在排序前,对浮游植物优势种丰度数据和除pH外的环境因子数据进行lg(x+1)转换。对浮游植物丰度数据采用去趋势分析(DCA),计算出DCA图中排序轴的最大长度,并据此进行单峰模型和线性模型的选取。采用蒙特卡洛置换检验筛选出影响浮游植物群落的关键环境变量。采用SPSS 24.0进行独立样本t检验。相似性百分比(SIMPER)分析和群落相似性分析(ANOSIM)采用Primer 5.0软件实现。
2 结果与分析
2.1 库里泡浮游植物群落分布特征
研究期间,共鉴定出浮游植物6门8纲13目23科49属106个分类单位。其中:绿藻门19属56种,占53%;硅藻门12属23种,占22%;其他门类占25%。库里泡浮游植物群落为绿藻-硅藻型。从时间上看,春季浮游植物种类数较低,以硅藻门、绿藻门和蓝藻门为主;夏季次之,主要为硅藻门、绿藻门和裸藻门;秋季最高,以硅藻门和绿藻门为主。从空间上看,浮游植物种类数呈现出湖泊区>入水口>出水口的分布状态。湖泊区浮游植物物种以蓝藻门、硅藻门和绿藻门为主,而出水口、入水口以绿藻门与硅藻门为主。库里泡浮游植物年均丰度为7.66×106cells/L,变化范围为4.97×106~10.89×106cells/L(图2)。浮游植物丰度在时间上呈现出秋季(3.63×106cells/L)>夏季(2.37×106cells/L)>春季(1.65×106cells/L)的分布趋势,且春、夏两季与秋季之间的浮游植物平均丰度差异显著(P<0.05);在空间上呈现出湖泊区(5.29×106cells/L)>出水口(1.28×106cells/L)>入水口(1.09×106cells/L)的分布趋势,且湖泊区与出水口、入水口之间的浮游植物平均丰度差异显著(P<0.05),但出水口与入水口之间的差异不显著(P>0.05)。

图2 库里泡浮游植物丰度和相对丰度的时空变化Fig.2 Spatio-temporal variation of phytoplankton abundanceand relative abundance in Kulipao Lake
研究期间,共鉴定出浮游植物优势种15种(表1)。从时间变化上看,除梅尼小环藻(Cyclotellameneghiniana)、狭形纤维藻(Ankistrodesmusangustus)、四尾栅藻(Scenedesmusquadricauda)为全年优势种外,优势种的演替特征是由春季的弱细颤藻(Oscillatoriatenuis)和短棘盘星藻(Pediastrumboryanum)演替为夏季的梅尼小环藻、绿色裸藻(Euglenaviridis)和裸甲藻(Gymnodiniumareuginosum),最后演替为秋季的狭形纤维藻和肥壮蹄形藻(Kirchneriellaobesa)。从空间变化上看,入水口的硅藻门优势种[如梅尼小环藻和谷皮菱形藻(Nitzschiapalea)等]的优势度远高于出水口,出水口的蓝藻门优势种(如弱细颤藻等)的优势度为全库里泡最高,而湖泊区的绿藻门和硅藻门主要优势种的种类数及优势度均高于出水口、入水口。

表1 库里泡不同采样区域和季节的浮游植物优势种及其优势度Table 1 Dominant species and dominance of phytoplankton in Kulipao Lake
ANOSIM结果显示(表2),时间变化对库里泡浮游植物群落的时空分布格局具有显著影响(P<0.05)。在空间上,出水口与入水口之间的浮游植物群落结构存在着显著差异(P<0.05),但湖泊区与出水口、入水口之间的差异并不显著(P>0.05)。SIMPER分析结果显示,梅尼小环藻、狭形纤维藻、弱细颤藻和绿色裸藻等是各采样区域的浮游植物群落结构产生差异的主要贡献物种,而导致浮游植物群落结构在时间上产生差异的主要贡献物种为梅尼小环藻、狭形纤维藻、弱细颤藻和球衣藻(Chlamydomonasglobosa)等。
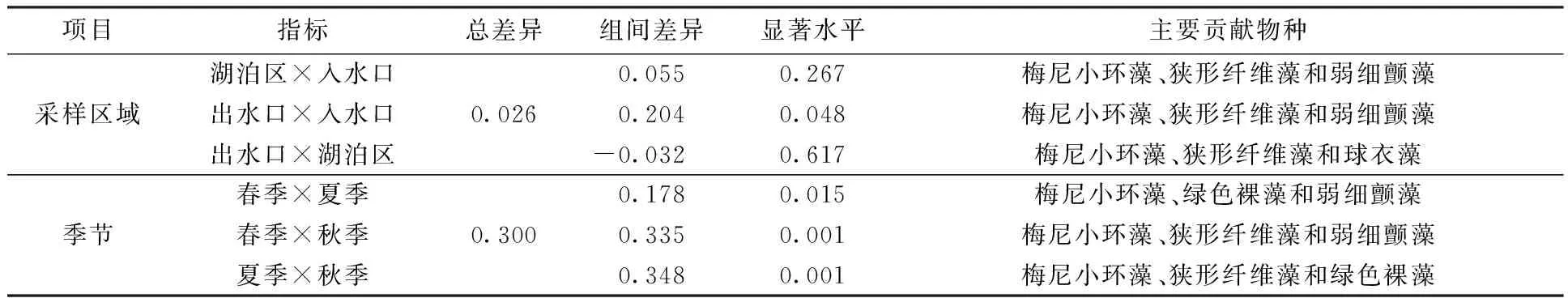
表2 库里泡浮游植物群落相似性分析及比较Table 2 Analysis and comparison of ANOSIM of phytoplankton community in Kulipao Lake
2.2 库里泡水体理化因子及营养状态指数
调查期间的水环境变量检测结果如表3和图3所示。研究期间,水体pH变化范围为8.23~9.04,均值为8.71,整体偏碱性;夏季水温(23.41 ℃)显著高于春(20.41 ℃)、秋(6.46 ℃)两季(P<0.05);电导率呈现春季最高、夏季最低的分布趋势,变化范围为621.50~2 282.63 μS/cm,均值为1 394.41 μS/cm;CODMn呈现秋季最高、夏季最低的变化趋势,变化范围为9.25~22.26 mg/L,均值为13.74 mg/L;TP呈现从春季到秋季逐步下降的变化趋势,变化范围为0.23~1.50 mg/L,均值为0.68 mg/L;DO和TN呈现随时间的推移逐渐升高的变化趋势,变化范围分别为6.00~1.34 mg/L和6.19~18.89 mg/L,均值分别为6.38 mg/L和0.79 mg/L;BOD5的年变化范围为0.26~2.21 mg/L,春(1.56 mg/L)、夏(1.55 mg/L)两季显著高于秋季(0.33 mg/L)(P<0.05)。独立样本t检验显示:水温、电导率、pH、BOD5的季节间差异显著(P<0.05);TP、TN和CODMn在季节间差异不显著,但在入水口与出水口之间差异显著(P<0.05);其他水环境指标(如pH、DO等)的空间分布态势变化不显著(P>0.05)。

表3 环境因子的平均值和变化范围Table 3 Averages and ranges of environmental variables
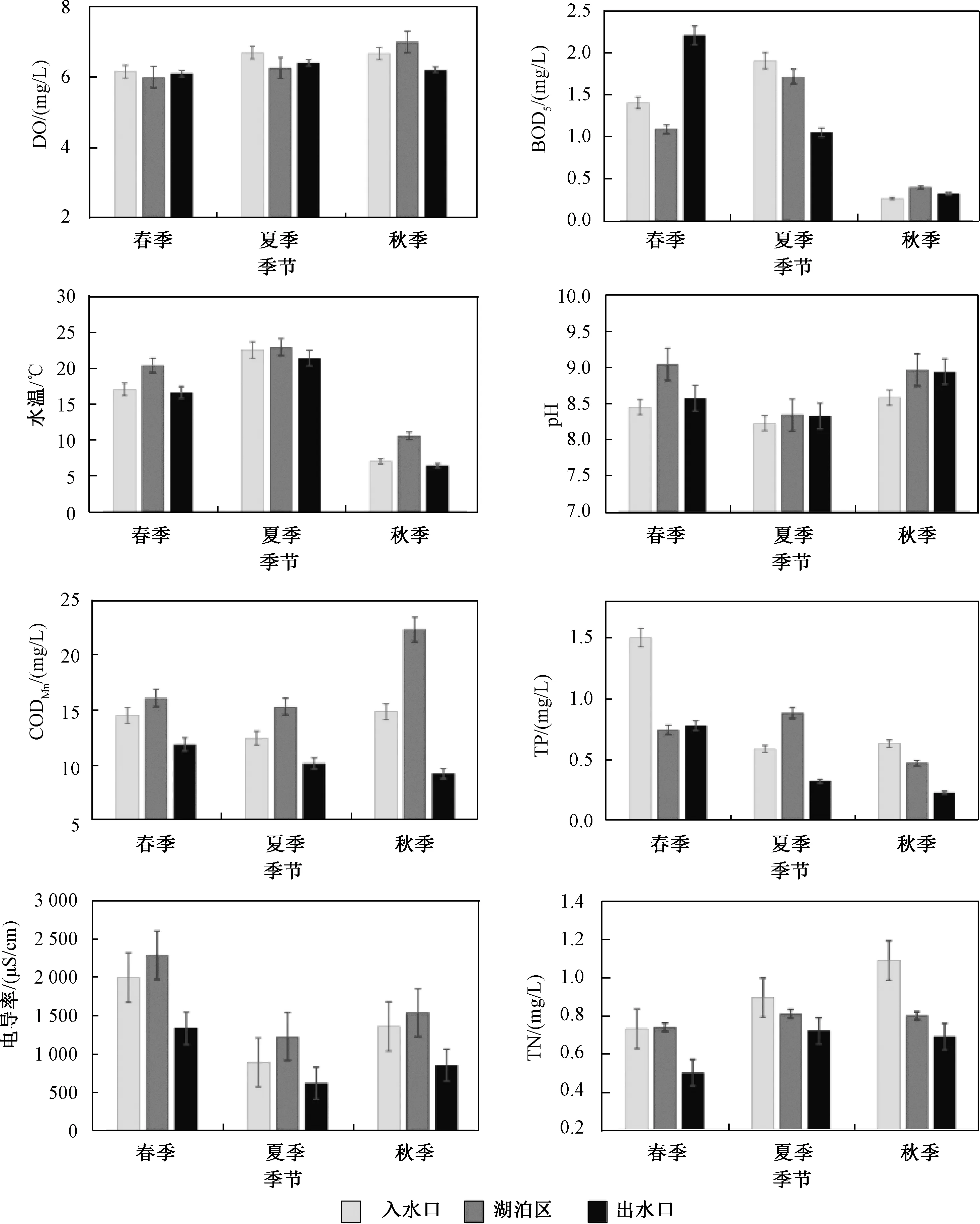
图3 环境因子的时空变化Fig.3 Spatio-temporal variation ofenvironmental variables
采用加权营养状态指数平均值对库里泡水体营养状态进行初步评价,结果如表4所示。研究期间,库里泡水体总体处于中度富营养状态。在空间上,入水口和湖泊区营养状态指数相对较高,出水口与入水口、湖泊区之间的营养状态指数差异显著(P<0.05);但在时间上,春、夏、秋3季之间的营养状态指数差异不显著(P>0.05)。
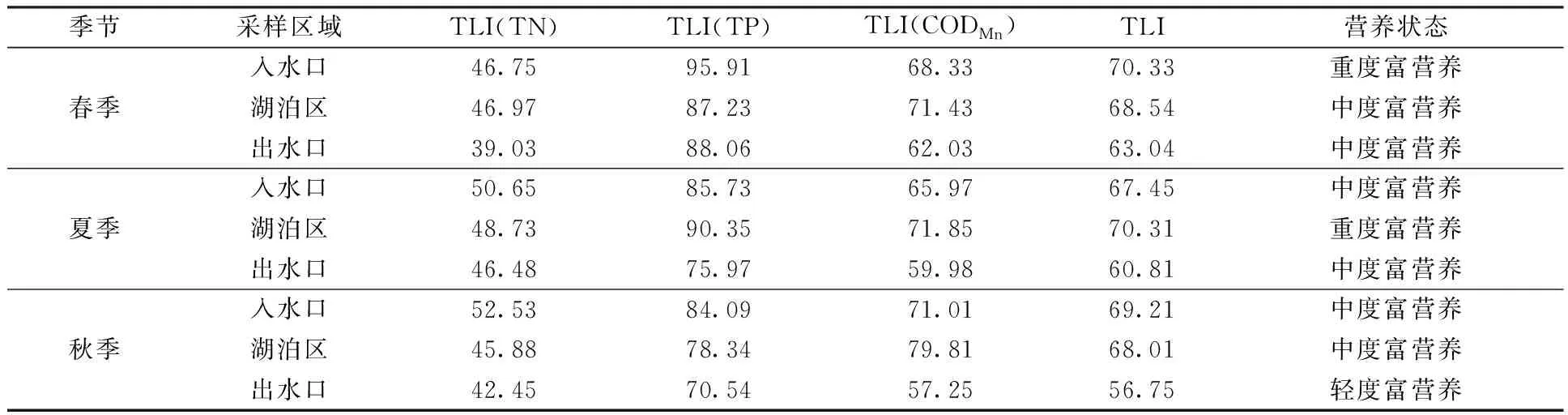
表4 营养状态指数评价结果Table 4 Results of TLI in Kulipao Lake
2.3 库里泡浮游植物多样性指数
应用Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数和Simpson生态优势度指数对库里泡浮游植物群落多样性进行分析,结果如图4所示。从空间分布来看,Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数和Simpson生态优势度指数的变化趋势均呈现为湖泊区>入水口>出水口。从时间分布来看,Margalef丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数均显示出明显的季节差异。丰富度指数表现为秋季>夏季>春季,其值随季节变化逐渐升高。均匀度指数和多样性指数的季节变化均呈现为夏季>春季>秋季。进入夏季后,多样性指数和均匀度指数逐渐升高至全年最高值;到了秋季,其数值逐步下降。Simpson生态优势度指数在各季节间的差异不显著。

图4 库里泡浮游植物多样性指数时空分布Fig.4 Spatio-temporal pattern of phytoplankton diversity index in Kulipao Lake
2.4 浮游植物群落与环境因子排序分析
对库里泡水温、TN、TP、BOD5、DO、电导率、pH和CODMn共8个环境变量与浮游植物优势种进行RDA,研究库里泡浮游植物群落的分布规律,并探究驱动浮游植物群落时空格局发生动态变化的关键环境变量。
DCA结果显示,排序轴最大长度梯度为1.717,表明RDA能够较好地阐明库里泡浮游植物的时空分布规律。RDA结果表明(图5),轴1、轴2的特征值分别为0.213、0.095,环境因子与浮游植物优势种的相关系数分别达到了0.890、0.814,前4个排序轴共同解释了88.9%的累积种类-环境因子百分比,基本反映了库里泡浮游植物群落时空演替情况及其与环境因子间的生态响应关系。经蒙特卡洛置换检验,水温(F=8.195,P=0.002)、TN(F=3.719,P=0.002)、TP(F=1.982,P=0.024)是驱动研究期间库里泡浮游植物群落结构发生时空变化的主要环境变量。

注:sp1,谷皮菱形藻;sp2,卵形双菱藻;sp3,梅尼小环藻;sp4,裸甲藻;sp5,卷曲鱼腥藻;sp6,绿色裸藻;sp7,湖生卵囊藻;sp8,短棘盘星藻;sp9,肥壮蹄形藻;sp10,狭形纤维藻;sp11,双对栅藻;sp12,四尾栅藻;sp13,球衣藻;sp14,卵形隐藻;sp15,弱细颤藻。图5 浮游植物种类、环境因子与样点间的RDA排序Fig.5 RDA ordination of phytoplankton species,environmental variables,and sampling sites
3 讨论
3.1 库里泡浮游植物群落结构变化特征
浮游植物群落分布特征与水环境因子之间关系密切。季节更替过程往往伴随着流域内水文条件的改变,浮游植物群落会通过调节更替机制来维持其生态系统的动态平衡,如浮游植物的资源(光、氮、磷、硅、碳等)获取能力和沉降速度等发生相应的改变[20-21]。在本研究中,库里泡浮游植物平均丰度呈现出夏季最低、秋季最高的变化趋势,这与天目湖[22]、镜泊湖[23]和Kinneret湖[24]所表现出的在温度较高的夏季的浮游植物丰度相对较高的状况明显不同。造成这一差异的主要原因可能是研究期间降雨量较大。研究表明,极端降雨会增加水体扰动,降低透明度和水温,进而导致水体浮游植物丰度降低[25]。2019年,进入8月份后,大庆市遭遇了近10年来罕见的暴雨,日均降雨量达120 mm。强降雨带来水体径流量的急剧增加及水体滞留时间的缩短,对研究区域浮游植物的生长代谢产生了一定程度的限制[26]。春、夏两季,库里泡蓝藻门的相对丰度较高,这与杨双[16]对库里泡浮游植物群落时空分布格局的研究结论类似。其原因与蓝藻自身的某些生理特性有关,如具有胶质鞘、气囊,以及抗拒紫外线伤害的能力,从而避免了春、夏两季强光照射等的影响,进而使之在水环境中处于优势地位[27-29]。此外,曹文钟等[7]在2012年5—10月对库里泡浮游植物进行了采样调查,发现其优势种主要为双对栅藻和梅尼小环藻,绿藻门和硅藻门优势种(如梅尼小环藻、四尾栅藻和狭形纤维藻等)的优势度在6—8月相对较高,与本研究对夏季浮游植物优势种种类及优势度的研究结果类似。本研究进一步表明,在以往的近10年里,库里泡浮游植物群落分布特征较为稳定。
近年来的浮游植物生态研究表明,硅藻门和绿藻门通常是我国温带湖库浮游植物群落的主要类群,在种类数量和丰度上都处于主要地位[10-15]。在本研究中,库里泡浮游植物的种类组成主要为绿藻-硅藻型。除在调查期间为全年优势种的狭形纤维藻、梅尼小环藻等种类外,春季的弱细颤藻、卷曲鱼腥藻和谷皮菱形藻等优势种会在夏季被绿色裸藻、裸甲藻等优势种取代,然后在秋季被肥壮蹄形藻和卵形双菱藻等优势种取代。库里泡水体氮磷比小于16∶1,氮限制明显。低氮含量能够显著制约浮游植物的发育,而蓝藻的繁殖积累与水体氮磷比息息相关。弱细颤藻和卷曲鱼腥藻等丝状蓝藻具备一定的固氮能力[29-30],而春季库里泡水体的氮水平相对较低,这可能就是弱细颤藻和卷曲鱼腥藻等蓝藻在春季占优势地位的主要原因之一。同时,SIMPER分析也表明,弱细颤藻是导致春季浮游植物群落结构与夏、秋两季相异的重要贡献种类之一。到了夏季,水温升高会导致喜温藻类植物的大量繁殖,表现为卵形隐藻、绿色裸藻和裸甲藻等偏爱高温的种群占据优势地位[31-33]。而进入秋季后,库里泡水体温度相对较低,光照强度相对较弱,部分长有鞭毛从而具有运动功能的绿藻可迅速占据生态空间,获得竞争优势[34-35],因此,球衣藻成为秋季的主要优势种。KATANO等[36]对低温环境下的衣藻生长状况的研究也得出了相同的结论,认为其原因在于衣藻碳富集机制的活性在低温下不会降低。此外,秋季库里泡水体中的氮含量相对较高,而狭形纤维藻是α~ms污染指示种,能够耐受低温环境且适宜在含氮量高的水体中生长[37],因此,狭形纤维藻在秋季占据一定优势地位。优势度较高的种类通常在群落中占据较宽的生态位,会降低物种间的资源竞争,最终导致群落稳定性降低,如有害藻华事件的发生[30]。有研究表明,优势种的变化对群落结构稳定性的影响是持续的,但是这种持续影响会受到环境因子的多重作用,导致不同环境条件下的优势种类趋于多元化[9,15]。
湖库对水体中营养盐的滞留作用能够显著降低流域内营养盐向下游的输出[38-39]。在本研究中,氮磷营养盐含量在入水口与出水口之间差异显著,可能是由于大庆市城市污水在通过安肇新河排入松花江前,会经历库里泡的泥沙过滤作用和生物过滤作用,使水体氮磷营养盐含量得到明显控制。ANOSIM结果显示,库里泡入水口和出水口在浮游植物群落结构上存在显著差异(P<0.05)。SIMPER分析表明,造成入水口和出水口浮游植物群落结构差异的重要贡献种类为狭形纤维藻、弱细颤藻和梅尼小环藻等。梅尼小环藻的比表面积大,能更高效地捕获水体中的氮磷营养盐[40];狭形纤维藻为淡水生境泛化种,可耐受高污性水体[37,41]。通常,高浓度的氮磷营养盐对于维持这两种藻类的持续生长有较高助益,可使之逐渐成为优势种群,因此,在本研究中,狭形纤维藻和梅尼小环藻能在营养盐含量更高的入水口占据优势地位,这与TILMAN等[42]对藻类植物半连续培养的研究结果一致。而弱细颤藻是一种藻丝直且呈胶质薄片状的漂浮群体,在流动水体中的分布较广泛,能通过运动寻找到适宜的环境,从而更好地吸收营养盐,实现快速生长繁殖[43]。鉴于狭形纤维藻、梅尼小环藻和弱细颤藻对污染水体具有一定的耐受性,三者的种群分布格局能在一定程度上表征库里泡环境变量的分布特征。库里泡物种多样性指数与综合营养状态指数均表现出湖泊区最高、出水口最低的分布趋势,这与原居林等[44]对南太湖的研究结果相近,即较高的营养盐含量促进了先锋种的定植,也促进了多样性指数的上升。本研究进一步验证了水体氮磷营养盐的空间分布格局能够对湖泊不同水文分区的浮游植物群落特征的形成产生重要的驱动作用。
3.2 浮游植物群落结构与环境因子的相关性
探索驱动浮游植物群落特征发生变化的关键水环境因子,是保护物种多样性和维持水生态系统稳定的切实有效手段[45]。大量研究表明,温带湖库浮游植物群落的演替主要受水温、光强、营养盐,以及原生动物、滤食性鱼类的牧食行为等上行和下行效应的影响[46-47]。在本研究中,水温、TN与TP是促进库里泡浮游植物群落演替的关键因子。RDA结果显示,浮游植物优势种的分布特征基本能够表征浮游植物群落对湖泊空间差异状况的生态响应关系。
水温是浮游植物季节间周期性变化的重要制约变量之一,可通过影响生物酶活性来调控浮游植物光合作用暗反应的速率,引起浮游植物生物合成与碳固定[48]。有研究表明,蓝藻是一种喜温性水生植物,水温对蓝藻发育、繁殖的助益效应十分显著[27,49]。这与研究期间库里泡春、夏两季蓝藻门丰度较高相符。结合RDA结果可知,在本研究中,水温在优势种弱细颤藻、卷曲鱼腥藻等蓝藻的春、夏两季爆发上起着至关重要的作用。同时,RDA结果还表明,绿藻门优势种四尾栅藻的丰度与水温呈显著正相关。郑忠明等[50]对不同温度下的四尾栅藻生长特性的研究表明,其适宜的生长温度为25~30 ℃。这一温度区间与本研究中春、夏两季的水温情况基本一致。因此,在春、夏两季,四尾栅藻能在与其他藻类植物的竞争中占据优势地位,继而成为优势种群。此外,有研究认为,水温上升可以大幅提升浮游植物的繁殖力[51],但在本研究期间,库里泡夏季降雨量大,水位不断上升,导致库里泡内的浮游植物难以正常繁殖积累[25]。因此,虽然库里泡夏季水温较高,但其浮游植物丰度却低于秋季。
TN是评估湖泊水体富营养状态的主要指标之一[52]。作为浮游植物繁殖代谢所必需的营养元素,当水体中的氮营养盐含量偏低或偏高时,往往会对浮游植物群落的分布格局构成影响[53]。DENG等[54]调查了连续8个季度的太湖监测数据后指出,TN、光强和TP等是影响太湖浮游植物群落分布特征的重要环境变量;王宇飞等[55]研究发现,汉丰湖浮游植物群落分布主要受TN、TP和水温等的影响。在本研究中,RDA结果显示,裸甲藻的相对丰度与水环境中的氮含量呈正向响应关系,夏季库里泡TN含量相对较高,裸甲藻的丰度也逐渐升高。这与CULLEN等[31]对美国加州La Jolla海湾裸甲藻水华的研究结果相近。CULLEN等[31]认为,这一现象不仅与裸甲藻的生理特征有关,还受到水生态系统的化学、物理和生物环境因素的影响。研究显示,裸甲藻对高氮废水的耐受性较高;在氮含量高而磷含量较低的水体中,裸甲藻相较于其他物种更具优势[56]。此外,有研究表明,水体氮磷比对藻类的暴发性生长存在重要影响;在较高氮磷比的时间段,绿藻的生长占优[57]。在本研究中,随着季节的推移,大庆市排向库里泡的工业污水中的氮含量呈现出逐渐升高的趋势,而磷含量则逐渐降低,因此,库里泡水体中的氮磷比呈现出随季节变化逐渐上升的趋势。这与库里泡绿藻丰度春季较低而秋季较高的现象相符合,也与BUZZI等[58]对意大利Como湖和吴文莉等[59]对处于富营养状态的我国安康水库的研究结果一致。
水体TP含量与浮游植物群落的时空分布关系紧密[60]。作为浮游植物物质生成和食物链能量流动的重要制约元素,水体TP含量的变化能够影响到浮游植物的生长发育及群落的动态分布[61]。在本研究中,大庆市排向库里泡的春季污水中的磷含量一直处于相对较高的水平,而秋季污水中的磷含量相对较低。库里泡春季出现的浮游植物以喜磷元素的种类为主,如短棘盘星藻和卷曲鱼腥藻等。RDA结果显示,短棘盘星藻与TP含量呈正相关,随着水体磷含量的升高,短棘盘星藻的丰度也随之升高。PALOHEIMO等[62]和贾竞宇等[63]也得到了相同的研究结果。这可能是由于低磷环境对短棘盘星藻的比增长率影响较大,随着磷质量浓度的增加,短棘盘星藻的生长作用会有一定的加强,进而可以形成较高的生物量,因此,春季水体环境可以为短棘盘星藻的生长提供充足的条件。卷曲鱼腥藻在库里泡春季水体中更具优势,而到了夏、秋两季则丧失了原有的优势地位。WAN等[64]对鱼腥藻生长特征的研究表明,鱼腥藻的生长对水体中的氮浓度的要求相对较低,但受磷浓度限制较显著。本研究结果与WAN等[64]的研究结论相似,即在磷源充足时,鱼腥藻易形成优势地位。这进一步印证了研究区域浮游植物优势种群与水体TP含量存在一定的响应关系,同时也说明水体中的磷含量对库里泡浮游植物群落特征的形成具有一定的驱动作用。孙玉凤[65]研究发现,2012年库里泡TP和TN含量的年平均值分别为1.44 mg/L和5.45 mg/L,相比入水口,出水口的TP和TN含量均较低,但差异不显著(P>0.05);杨双[16]通过水质调查发现,2014—2015年库里泡TP和TN含量的年平均值分别为1.18、1.92 mg/L和3.77、2.32 mg/L,两年间的出水口、入水口TP和TN含量差异均不显著(P>0.05)。与上述研究相比,在本研究期间,库里泡TP和TN含量均相对较低,年平均值分别为0.68 mg/L和0.79 mg/L,出水口TN含量显著低于入水口(P<0.05),表明库里泡水体中的TP和TN含量呈逐年降低趋势,湖泊区水体自净能力较之前有一定程度的提升,水质状况总体有所改善。在未来人为活动强度不变的情况下,持续关注库里泡水体营养状态是加强水生生物多样性保护的关键。
整体来看,库里泡作为一个受季风影响的温带富营养化中小型湖泊,其水温存在显著的季节性变化,且氮磷营养盐浓度常年较高,近10年间的浮游植物群落结构相对稳定。在本研究中,水温及氮磷营养盐含量成为影响库里泡浮游植物群落动态分布格局的重要环境变量,这也反映了典型的温带地区中小型城市富营养化湖泊浮游植物的生态学特征。
4 结论
1)研究期间,本研究在库里泡共鉴定出浮游植物106个分类单位。库里泡浮游植物群落组成为绿藻-硅藻型,全年优势种主要为梅尼小环藻、狭形纤维藻和四尾栅藻等。
2)库里泡浮游植物群落时空变化显著。在时间上,浮游植物丰度呈现秋季>夏季>春季的变化趋势。在空间上,入水口的浮游植物群落以硅藻门、绿藻门和蓝藻门为主,出水口以绿藻门、硅藻门和隐藻门为主,湖泊区浮游植物丰度显著高于出水口、入水口。
3)浮游植物群落对库里泡水环境因子时空异质性的动态响应较为明显,水温、TP和TN是影响库里泡浮游植物优势种组成的关键环境变量。
4)库里泡水体整体处于中度富营养状态。与近10年间的库里泡相关研究进行对比,发现其当前的水体富营养状况有所改善。库里泡浮游植物丰度和氮磷营养盐含量均为入水口较高、出水口较低,说明湖泊区对氮磷营养盐有一定的过滤作用,进而对浮游植物群落结构产生了一定影响。
猜你喜欢
当代水产(2022年6期)2022-06-29
环境工程技术学报(2022年3期)2022-06-05
流行色(2019年10期)2019-12-06
装饰装修天地(2019年9期)2019-10-21
南水北调与水利科技(2019年2期)2019-06-24
陕西档案(2019年2期)2019-05-07
绿色科技(2016年24期)2017-03-30
河北渔业(2017年1期)2017-02-21
当代工人(2016年7期)2016-05-18
小雪花·成长指南(2015年10期)2015-10-23
