
内生菌定殖、促生及降低作物有机污染的机理研究(Ⅰ):定殖与促生
2022-09-02 02:22张妃冯乃宪宾慧李彦文赵海明向垒蔡全英莫测辉
农业环境科学学报 2022年8期
张妃,冯乃宪,宾慧,李彦文,赵海明,向垒,蔡全英,莫测辉
(暨南大学生命科学技术学院,广东省环境污染控制与修复材料工程中心,广州 510632)
内生菌是指在其生活史某一阶段或整个阶段生活在植物体内,且不会引发植物明显病害症状,与植物互利共生的一类微生物。内生菌通过植物根系皮层伤口或缝隙和叶片气孔、水孔等进入植物体内,然后通过维管系统迁移至其他部位,并定殖于根、茎、叶、花、果实等器官中。目前,已报道的内生菌有100 余种,隶属于50 余个菌属,主要包括芽孢菌属()、假单胞菌属()、伯克霍尔德氏菌属()等(图1)。内生菌栖息于植物体内并与宿主植物协同进化,形成了植物生态系统中最古老、最广泛的互惠共生关系。内生菌识别宿主植物并定殖于植物体内的过程主要受到菌株自身遗传特性、植物类型和品种、根系分泌物化学成分,以及土壤理化性质等因素的影响。
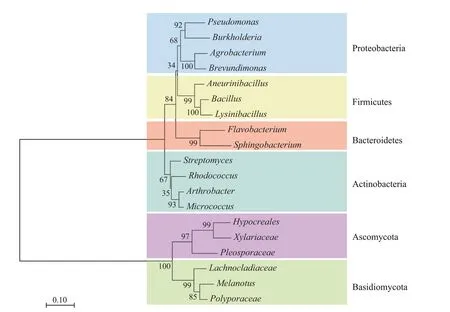
图1 来源于不同植物的部分内生菌的系统进化树Figure 1 Phylogenetic tree of different groups of endophytes isolated from different plants
内生菌是植物微生态系统的重要组成成员,被称为植物“第二基因组”,内生菌通过自身代谢活性可直接或间接地促进植物养分吸收,调节植物生理过程,维持植物体内微生态平衡,保障植物健康生长,而宿主植物则会为内生菌生存提供稳定的空间和丰富的碳源。近年来,有关内生菌在植物微生态调控、根际或叶际免疫、逆境适应、养分吸收、碳循环、多样性维持等方面的研究取得了一系列重要进展。已有的有关内生菌的综述论文只涉及了定殖、来源、促生作用、重金属和有机污染物修复等某个或某几个方面,但缺乏对这些研究系统整体的认识,而且对其中相关研究的分子机制了解也不够深入。为此,本研究针对内生菌定殖、促生作用和降低作物体内有机污染物的机理进行系列综述,其中包括、、、等期刊具有代表性的最新研究成果,为推动内生菌与植物相互作用研究的发展、促进土壤污染修复和保障农产品安全提供理论依据。本文属于该系列综述的第一篇,着重阐明内生菌的来源、与宿主植物的识别机制和定殖途径、定殖的影响因素和促生作用机制,并指出目前研究中存在的问题和未来的研究方向;内生菌降低作物体内有机污染物的相关内容将在《内生菌定殖、促生及降低作物有机污染物含量的机理研究(Ⅱ):降低作物有机污染机理》中进行详细论述。
1 内生菌的定殖
1.1 内生菌的来源与定殖途径
种子是植物繁殖的重要器官,其内部栖息着种类丰富且结构复杂的微生物群落,因此也是内生菌通过垂直传播方式在连续的植物世代之间传递的载体,这种传播方式提高了从母本选择性传递有益内生菌的概率,从而确保内生菌在子代植物体内稳定存在,直接或间接影响种子活力和幼苗成活率。种子内生菌是子代植物内生菌群建立的基础,可通过快速占据子代植物有利的生态位,进一步影响子代植株内生菌群的装配和微生态的动态平衡。水稻亲本对子代种子内生菌群的建成具有重要影响,通过杂交、近亲繁殖和人工选择后的水稻种子中具有的相似内生菌种群可作为核心菌群跨代传播。目前已报道的种子内生细菌主要分布于变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidetes),种子内生真菌主要分布于子囊菌门(Ascomycota)和担子菌门(Basidiomycota)。种子内生菌长期定殖于种子内部,是宿主植物的延伸免疫系统,其作为抗性的传导剂可提高子代植物抗病性和环境适应性。如在抗病表型的水稻种子中世代积累和传播的内生细菌鞘氨醇单胞菌()可通过产生抗病性信号分子邻氨基苯甲酸来干扰病原菌伯克霍尔德氏菌()的sigma 转录起始因子RpoS,从而抑制病原菌致病因子的生物合成,赋予易感表型的水稻品种对该病原菌的抗性。
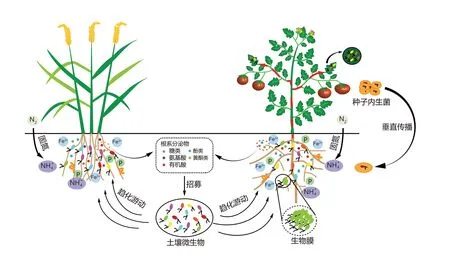
根际土壤中含有高度多样化的微生物群落,其菌落总数高达10~10CFU·g(土壤湿质量),因此也是内生菌的重要来源。扩增选择模型显示,根际微生物被宿主植物选择性招募,对根系分泌物产生趋化作用,向植物根际、根表游动,然后在根表形成微菌落、微基团、共质体或生物膜等,借助根系裂缝、伤口、根毛等孔口进入植物体内,最后在植物组织内部扩增繁殖,使植物组织中内生菌数量达到10~10CFU·g(鲜质量)(图2)。例如,固氮根瘤菌()利用侧根基部及皮层细胞间的裂缝进入植物体内定殖。某些根际微生物分泌的细胞壁水解酶可破坏植物细胞壁,突破细胞壁的门控作用,依次穿过皮层、内皮层和中柱,进入木质部导管,最后在鞭毛运动和植物蒸腾拉力协同作用下穿越植物的多个生理屏障,从而实现由根部向地上部的迁移和定殖,如葡萄根际的伯克霍尔德氏菌(PsJN)可进入根系并迁移至花序组织中定殖。研究表明,在植物木质部管腔定殖是植物病原菌的特性,而内生菌只定殖在植物导管内部或通过质外体在植物体内迁移,但也有研究发现内生菌可以在木质部管腔内定殖。一些根部的内生真菌可依靠菌丝沿着宿主植物的木质部导管腔扩散到整株植物。

图2 内生菌的定殖及促生作用机制[1,6]Figure 2 The colonization and plant growth-promoting mechanisms of endophytes[1,6]
植物叶片表面含有多种微生物类群,它们通常附生于叶片,通过气孔、排水器、伤口等进入叶片质外体,然后迁移到内皮层细胞中生长繁殖,并可向下迁移至植物茎干和根系。叶片表面附生的微生物进入叶内组织不完全是被动过程,它们可以分泌一些低于植物病原体表达浓度的细胞壁水解酶,从而主动地进入叶片组织。宿主植物通过其叶片表面不同的化学和物理特征,并通过免疫活性、分子信号传导和屏障形成等对叶片内生菌的丰度和群落组成进行控制。研究指出,宿主植物的邻域扩散效应可塑造叶际微生物的多样性,因此,宿主植物的品种和叶龄、邻近植物的种类和生物量或年龄等生物因素,以及强光、高温和紫外线等非生物因素均是影响叶片内生菌定殖的重要因素。
1.2 影响内生菌定殖的关键因素
内生菌与宿主植物协同进化,具备了成功侵入和定殖于宿主植物体内的能力。内生菌成功定殖需同时具备3 个条件:(1)具有一定的黏附力,形成微群落或生物膜,黏附在植物表层;(2)适宜的栖息环境,保障微生物的生存和繁殖;(3)具有群体感应,保证足够数量的微生物细胞。内生菌定殖于植物体受多种因素的影响,主要包括内生菌自身遗传特性、植物类型和品种、根系分泌物,以及土壤理化性质等。
1.2.1 内生菌自身遗传特性
内生菌的自身遗传特性如鞭毛、菌毛、趋化蛋白、细胞壁溶解酶、胞外聚合物以及根瘤菌的结瘤因子等影响其在植物体内的定殖能力。鞭毛游动驱动菌体细胞对宿主植物的根系分泌物和根部黏液产生趋化作用,使内生菌向根部聚集并形成根表生物膜,随后进入植物体内定殖。内生菌沙雷氏菌(sp.S119)和肠杆菌(sp.J49)借助鞭毛运动,以泳动、群游和蹭动等方式向玉米、花生和大豆的根系趋化游动,形成根表生物膜,然后定殖于植物体内;内生固氮弧菌(sp.CIB)体内的Ⅳ型鞭毛和菌毛可驱动菌体细胞的运动和在水稻体内的定殖,若鞭毛蛋白基因发生突变,则会导致其在宿主水稻体内定殖的数量降低一个数量级。位于细胞表面的趋化受体蛋白(MCP)可介导内生菌对根际趋化物的感知,调控内生菌向根表趋化游动和定殖,例如:内生菌枯草芽孢杆菌(NCIB3610)的趋化受体蛋白McpB、McpC 和TlpC 可识别拟南芥()根系分泌物中的信号分子,诱导该菌株高效定殖于植物体内;内生菌阴沟肠杆菌(Ghats1)基因组中含有趋化受体蛋白基因(、、等)和鞭毛合成与组装基因,这些功能基因与该菌株定殖及适应植物体内环境密切相关。
内生菌借助微生物细胞之间的群体感应机制感知自身或其他微生物细胞密度和周围环境变化,使内生菌群体实现个体无法完成的在植物体内的定殖行为和生理功能。内生菌慢生根瘤菌(sp.ORS278)中LuxI-LuxR 型群体感应系统产生的肉桂酰-高丝氨酸内酯(HSL)信号分子可参与调控菌株ORS278 生物膜的合成和细胞运动,提高其在水稻根部的定殖效率。科萨克氏菌(sp.KO348)中的PsrR 群体感应系统参与了该菌株在水稻根内的定殖过程。然而,某些内生菌在植物体内的定殖过程并不受群体感应系统的调控,如群体感应信号分子N-酰基高丝氨酸内酯(AHLs)与内生菌沙雷氏菌(sp.)在番茄和油菜根部的定殖行为之间不存在显著相关性。
1.2.2 植物类型和品种
内生菌在植物体内的定殖过程符合扩增选择模型,根际微生物在根际大量繁殖扩增,受到宿主植物的选择性招募后,逐渐向多样性更低、特化程度更高的方向发展,最终被装配到宿主植物体内,如联合固氮的内生菌在甘蔗、水稻和玉米等禾本科植物的根、茎和叶内定殖,而共生固氮的根瘤菌在花生、大豆等豆科根部定殖。植物类型影响内生菌的装配和定殖,如沙鞭()、羊柴()和黑沙蒿()3 种植物的内生菌群组成和结构变异度高达47%。降解石油烃的内生菌不动杆菌(sp.BRSI56)和绿脓假单胞菌(BRRI54)在巴拉草()体内的定殖效率与代谢活性均高于双稃草(),从而使巴拉草对石油污染的修复能力也强于双稃草。重金属Pb/Zn 超富集植物小花南芥()和土荆芥()根和茎内的优势菌为假单胞菌(),其丰度占总内生菌类群的41.6%~81.56%,但其内生细菌多样性指数(Chao 1 和Shannon 指数)显著低于非Pb/Zn 超富集植物纤毛婆婆纳()。
植物品种(基因型)差异同样影响内生菌的定殖,如抗草甘膦转基因大豆()根部内生菌的多样性和丰度高于野生型大豆;高淀粉基因型甘薯()根部的优势内生菌为厚壁菌门的芽孢杆菌属,低淀粉基因型甘薯根部的优势内生菌为变形菌门。植物组织类型会对内生菌的种类、数量产生较大影响,如小麦()根部内生菌的种类比叶内更丰富,且根和叶内的优势菌种类完全不同;而沙漠灌木茎部的内生真菌多样性和群落组成的变异度却显著高于叶和根。
1.2.3 根系分泌物
根系分泌物中含有的特定信号物质(包括氨基酸、低分子有机酸、糖类、酚类等)能够影响内生菌的定殖。根际微生物通过感受根系分泌的信号物质向根表游动并形成稳定的生物膜,最后在植物体内定殖。如粪产碱杆菌()对秋葵()根系分泌物中的酚酸类物质(莽草酸、延胡索酸、香草酸和槲皮素等)产生趋化作用,向根表游动并形成生物膜,进而定殖在植物体内。拟南芥通过调节体内水杨酸和茉莉酸合成基因的表达,改变根系分泌物中水杨酸和茉莉酸的浓度,进而调控其内生菌群的装配过程。玉米根系分泌物中的黏液物质(如根尖脱落的水合多糖)可诱导固氮螺菌(Sp7)在根内的定殖。根系分泌物中的糖类物质如蔗糖能够触发枯草芽孢杆菌()胞外多聚胆固醇合成的级联反应,诱导菌体产生大量表面活性物质和超鞭毛,从而促进杆菌和假单胞菌在拟南芥根部及体内的定殖。黄瓜根系分泌物中的D-半乳糖诱导贝莱斯芽孢杆菌(SQR9)向根系游动并形成生物膜,促进该菌株的根系定殖。
根系分泌物中的黄酮类物质是目前研究较为深入的信号物质。玉米根系分泌物中的木犀草素、毛异黄酮、酚类咖啡酸等黄酮类物质可诱导内生菌红绶曲霉(wlg2)在根系的定殖。黄酮类物质也是根瘤菌定殖的关键信号物质。大豆、花生、豌豆等豆科植物根系释放的类黄酮/异黄酮、大豆苷元等物质可促进根瘤菌的定殖和结瘤。花生根系分泌物中的槲皮素、异甘草素和鹰嘴豆芽素A 等黄酮类物质能够提高根瘤菌(sp.CCTCCAB93190)结瘤基因的表达量,使结瘤数显著增加。菜豆()和百脉根()中的类黄酮芹菜素能够诱导根瘤菌(CIAT 899)中结瘤基因和的表达量显著上调,从而促进该菌株在根系中的高效定殖。
1.2.4 土壤理化性质
土壤理化性质如pH、有机质含量、孔隙度等显著影响植物根系的生理活动和根系分泌物组分,进而影响内生菌在植物根系的定殖。通常情况下内生菌在高孔隙度土壤中的植物体内的定殖效率更高;内生菌在高有机质含量土壤中的植物体内的多样性更高,同种土壤类型中的不同植物体内具有相似的内生菌群。此外,研究发现内生菌在沙鞭和羊柴浅层土壤(0~30 cm)根系中的总定殖率显著高于深层土壤(30~50 cm)根系;连续耕作5 年以上的农田中玉米内生细菌的相对丰度(47 个菌属)高于休耕5 年以上农田中玉米内生细菌的丰度(30个菌属)。
2 内生菌的促生作用
2.1 固氮作用
植物所需氮元素的60%~80%来源于微生物的固氮作用。许多内生菌具有固氮活性,可将大气中的N转化为NO、NH等,从而为宿主植物的生长发育提供丰富的氮元素。根据内生固氮菌与宿主植物的关系,将内生固氮菌分为专性内生固氮菌和兼性内生固氮菌。专性内生固氮菌包括根瘤菌()、固氮螺菌()等,其宿主主要是豆科植物;兼性内生固氮菌包括假单胞菌()、伯克霍尔德氏菌()、肠杆菌()和芽孢杆菌()等,它们与甘蔗、水稻等禾本科植物联合固氮。研究发现,接种内生固氮菌施氏假单胞菌(A15)能显著促进水稻幼苗生长,促生效果优于化肥。内生固氮菌多黏类芽孢杆菌(P2b-2R)定殖于油菜体内,可为宿主提供所需氮元素,促进植株的生长发育。水稻接种共生的丛枝菌根真菌根内球囊霉()可诱导根系硝酸盐转运基因4.5 超表达,提高水稻根系对氮素的吸收效率,基因4.5 缺失会导致氮素吸收率显著降低以及定殖效率下降。近期研究发现,生活在贫营养地中海的海草()根组织中的内生固氮菌与海草形成共生关系,内生固氮菌为海草提供氨和氨基酸,海草为内生固氮菌提供糖类物质。内生固氮菌与宿主植物建立了密切的共生关系,为植物提供了丰富的氮元素。对非豆科植物来说,内生固氮菌将成为农业生产中潜在的稳定氮源。内生固氮菌种类繁多、固氮类型和途径多样,因此内生固氮菌与宿主植物间的分子识别及相关固氮分子机制亟待深入研究。
2.2 溶磷作用
土壤中约95%的磷为无效态,无法被植物直接吸收利用。内生菌通过自身代谢产物将土壤中无效态磷活化为能被植物直接吸收的有效态磷。目前已报道的溶磷内生菌主要分布于放线菌门、厚壁菌门和变形菌门,隶属于17 个菌属。根际酸化为内生菌溶磷的主要机制之一,内生菌分泌的大量有机酸(如柠檬酸、甲酸、葡萄糖酸、苹果酸和草酸等)可降低根际土壤pH,将磷酸钙、磷酸铁等难溶性磷酸盐溶解为可溶性的单碱性(HPO)和双碱性磷酸根离子(HPO),或者利用有机酸络合Ca、Fe等阳离子,释放PO。葡萄糖酸(GA)的产生是溶磷菌解磷的重要机制。GA 脱氢酶(GDH)是产生GA 的关键酶,GDH 的辅酶吡咯喹啉醌(PQQ)是GA 产生的关键因子,水稻内生菌草螺菌(Z67)异源表达PQQ 合成基因和GDH 合成基因,产生的PQQ 和GA 显著增加了宿主植物水稻体内的磷含量。编码GDH 的另一个功能基因也参与GA 的合成,增强溶磷活性,如多株内生菌荧光假单胞菌(L111、L228和L321)携带的基因均具有较高的表达活性,能够产生较多的GA(4 940~33 240 mg·L),具有较强的溶磷活性(438~1 312 mg·L),可显著增加宿主植物豌豆的生物量。
除根际酸化外,内生菌分泌的植酸酶、C-P 裂解酶和核酸酶等可将有机质中的有机磷(如磷酸酐或磷脂)脱磷酸化,释放磷元素,如内生菌铜绿假单胞菌(BAC3115)、嗜麦芽寡养单胞菌(BAC2135)、微杆菌(BAC1157)和红平红球菌(BAC2162)均具有较高的植酸酶活性,其水解肌醇六磷酸中的磷酸单酯键,可释放肌醇磷酸酯和无机磷酸。目前,国内外学者对内生菌溶磷能力和机制进行了较多研究,但总体上还停留在表观溶磷活性变化和个别有机酸溶磷机制等方面,仍不成体系。溶磷机制复杂多样,将来应着重采用多组学和稳定同位素探针(如RNA-SIP、DNA-SIP)等技术研究内生菌溶磷的分子机制。
2.3 产铁载体
土壤中的铁元素主要以微溶形式存在,不易被植物吸收利用。限铁条件下,内生菌产生铁载体(又称嗜铁素),从其他化合物中抢夺Fe,或者将土壤中难溶性铁转化为可溶性铁,从而缓解宿主植物缺铁状况。铁载体是一类重要的生物活性物质,能够高效且特异性地螯合Fe。根据铁载体螯合基团的化学特性,可将其分为氧肟酸类、儿茶酚类和混合型(也称柠檬酸型)3 种类型。内生菌中的假单胞菌属是铁载体的高产菌株,产生的铜绿假单胞菌铁载体、绿脓杆菌螯铁蛋白、亚铁氰化铁等多种铁载体,可为宿主植物提供丰富的铁元素。内生菌产生的铁载体与Fe的螯合物可被植物直接吸收利用,这可能是植物抗缺铁胁迫的主要机理之一。目前,关于内生菌分泌铁载体在植物铁营养调控作用中的分子机制尚不明确,还亟待深入研究。
内生菌分泌铁载体也是提高宿主植物抵御病原菌的一种重要机制,内生菌通过铁载体与病原菌竞争有限的Fe,抢占生态位,使病原菌缺Fe失活而达到抑制病原菌的目的,如胡椒内生菌恶臭假单胞菌(Pt12)产生的铁载体可抑制病原菌镰孢霉菌(sp.)。目前,关于铁载体的研究主要在实验室内进行,实际环境中内生菌产生的铁载体的作用还与Fe浓度、内生菌活性、土壤理化性质等密切相关,这些还亟待开展专题深入研究。
2.4 分泌植物激素
内生菌自身代谢产生或强化宿主植物合成的生长素吲哚乙酸(IAA)、赤霉素(GAs)、细胞分裂素(CTK)等植物激素,可以作为介导植物生理过程的关键内源因子,调控植物生长发育和胁迫响应。内生菌合成IAA,改变植物内源IAA 含量,调节植物的生理与发育过程。如产IAA的内生菌能够增加玉米种子的IAA含量,使玉米幼苗根系分泌物中的IAA 浓度显著增加。产IAA 的内生菌教酒链霉菌(WZS021)能够显著增加宿主植物甘蔗体内IAA的含量,促进根系发育和生物量增加,提高抵御干旱胁迫的能力。此外,内生菌也可以降解IAA,消除过高浓度IAA 对植物生长的抑制作用,如内生菌恶臭假单胞菌(1290)能够降解植物体内过量的IAA,维持宿主植物体内IAA 的动态平衡,消除IAA 对萝卜根伸长的抑制作用。一些内生菌能够产生GAs,调控植物生长发育,提高宿主植物抵御各种逆境胁迫的能力。如内生菌烟曲霉(TS1)和层生镰刀菌(BRL1)可分泌多种GAs 和IAA,提高宿主水稻的叶绿素含量和生物量;内生菌假单胞菌()能够分泌GAs和IAA,促进胡萝卜块根的形成。内生菌甲基杆菌(sp.)能够分泌CTK,显著提高小扁豆()的光合速率和茎、根的生物量。
乙烯是植物在抵抗逆境生理过程中应激产生的植物激素。某些内生菌分泌的1-氨基环丙烷-1羧酸(ACC)脱氨酶,可将合成乙烯的直接前体物ACC分解为α-丁酮酸和氨,降低植物体内乙烯含量,维持逆境胁迫下植物体内乙烯的适量水平,保证植物正常生长。内生菌假单胞菌(8R6)能够合成ACC 脱氨酶,调控长春花体内乙烯的含量,增强对植物病原菌的抗性。内生菌枯草芽孢杆菌和荧光假单胞菌携带的ACC 脱氨酶,能够显著降低宿主植物豌豆()体内ACC 合成酶和ACC 氧化酶的活性,降低乙烯含量,增强豌豆对盐胁迫的抗性。将外源ACC 脱氨酶编码基因导入内生菌格氏沙雷菌(BXF1)体内进行异源超表达,定殖后能够促进宿主植物大豆的生长并抑制植物病原菌。不同植物激素相互耦联且能调控广泛的生理过程,从而形成复杂的植物激素调控网络,未来应重点研究内生菌对植物激素调控网络的调节作用。
3 展望
内生菌定殖在宿主植物体内,建立了密切的互利共生关系。研究内生菌定殖和促生机制,揭示内生菌-植物之间的分子识别机制,充分挖掘内生菌-植物构成的微生态系统的整体功能,对保障化肥农药减施增效和绿色农业可持续发展具有重要意义。本文重点梳理了内生菌来源、定殖和促生作用机制方面的最新研究进展。然而,我们仍然缺乏对内生菌-植物互作机制的系统认识。基于植物-微生物组构成的生命共同体,利用多组学、基因编辑和探针标记等先进技术,阐明内生菌-植物关键界面的互作机制,可以从以下方面深入研究:
(1)以往内生菌研究多采用常规的组织分离方法,获得的多是好氧型、生长快速的菌株,一些厌氧型、生长缓慢或不可培养的内生菌易被遗漏,难以满足目前对内生菌资源挖掘的需求。未来可从改善菌株分离培养方法入手,利用培养组学技术探索内生菌原位培养策略,利用单细胞基因组学和DNA-SIP 追踪内生菌和植物互作过程中同位素(如C-、N-或O-等)标记的化合物代谢途径,鉴定参与物质代谢的关键菌株,这将有望解决某些内生菌群难培养甚至不可培养的难题。
(2)内生菌与植物互作关系十分复杂,内生菌与宿主植物之间的信号识别机制不是依靠单个或一组基因控制的。目前针对内生菌与植物互作关系的研究主要关注单个时间点内生菌或植物的响应,未涉及内生菌与植物之间动态互作关系。因此,未来可从植物、根(叶)际微生物组、土壤等不同角度定向突破内生菌-植物互作的难题,以模式植物(如拟南芥)、模式菌株(如枯草芽孢杆菌)为切入点,采用多组学和CRISPR-Cas9 基因编辑等先进技术全面挖掘参与内生菌定殖的功能基因和信号分子,实时监控内生菌-植物间的信号网络和动态调控机制。
(3)目前,内生菌与植物互作研究主要针对单一微生物,功能较单一,且易受环境因素影响。因此,未来可利用培养组学和代谢组学技术,结合数学模型,研究核心内生菌群落的定殖和装配机制,揭示核心内生菌群与植物互作的分子机制,进而选择最佳接种菌剂、接种时机和部位,以最大程度地提高内生菌群的生态服务功能。
猜你喜欢
中国德育(2022年18期)2022-10-14
伴侣(2022年5期)2022-06-16
读者·校园版(2020年7期)2020-04-09
湖北农业科学(2019年22期)2019-12-23
农家致富顾问·下半月(2019年7期)2019-09-28
新媒体研究(2018年6期)2018-07-03
大众科学(2018年1期)2018-03-28
科教导刊(2017年6期)2017-03-27
小星星·阅读100分(高年级)(2015年11期)2015-11-28
大自然探索(2015年11期)2015-09-10
