
北京山区侧柏林下草本植物多样性的影响因素分析
2022-08-31 03:33:04伍冰晨崔冉冉
草地学报 2022年8期
张 林, 齐 实*, 周 飘, 伍冰晨, 张 岱, 崔冉冉, 黄 宪, 马 宁
(1.北京林业大学水土保持学院, 水土保持国家林业局重点实验室, 北京 100083)
林下草本植物作为森林群落的重要组成部分,在提升土壤质量和维持生态系统稳定等方面发挥着不可替代的作用[1-4]。但林下草本植物的组成和分布受林分结构、土壤和立地条件等多因素综合影响[5-10],因此解析林下草本植物多样性的维持机制对森林的可持续经营具有重要的理论指导意义[11]。
近年来,研究倾向于分析气候、地形、土壤等[12-14]环境因子对林下植物多样性的影响,常按降雨、海拔、坡向、土壤等[15-17]环境因子给予定性或定量描述,再对比分析不同因子下的植物多样性差异。王子婷等[18]研究发现,坡向和土壤水分是限制黄土区坡面草本植物多样性的主导因子;温佩颖等[19]认为林下草本物种多样性主要受海拔和凹凸度的影响;李婷婷等[20]指出,土壤有机质和pH是影响柏木(Cupressusfunebris)林下草本物种多样性的主要土壤因子。也有学者认为,林下草本物种多样性的差异不仅归因于环境因子,还受到上层林分结构的显著影响[21-22],但对林下草本物种多样性的作用结果缺乏共识,陈丝露等[23]以不同改造模式的柏木林为研究对象,认为混交林改造模式对林下草本物种多样性具有抑制作用;而曹小玉等[24]认为混交度对林下草本物种多样性具有促进作用;张柳桦等[25]以马尾松(Pinusmassoniana)林为研究对象,认为低密度对林下物种多样性具有促进作用;而张勇强等[26]认为中等林分密度最适合林下草本植物的生长和发育。林分结构与环境因子有着密切的联系,海拔梯度和土壤养分的变化在区域范围内被认为是植物生长发育的主要因素,同时,上层林分结构对土壤理化性质具有显著影响,两者共同影响着林下草本物种多样性[27-28]。仅将林下草本物种多样性的差异归因于单个林分结构因子或环境因子,而忽略由环境因子和林分结构因子共同形成的耦合影响,难以充分揭示影响林下草本植物多样性的真实机理。此外,多数研究倾向于采用传统的线性回归分析方法,而忽略了各影响因子之间的交互作用。因此,在对林下草本物种多样性的影响因素研究中,需采取科学合理的统计方法来综合分析环境因子和林分结构因子对林下草本物种多样性的相对贡献。结构方程模型(Structural equation model,SEM)是一种基于验证性因素分析和路径分析来揭示某一现象的统计方法,对于模拟和验证多因子间的复杂关系具有巨大的优势[29],从而可以更加全面的揭示林分结构因子和环境因子对林下物种多样性的耦合影响机制。
侧柏(Platycladusorientalis)是北京市京津风沙源二期林业工程的主要造林树种,工程目的在于改善林分结构,提高林下物种多样性。因此,如何定量分析林分结构因子和环境因子对侧柏林下物种多样性的影响,并采取有效措施提高其林下植物多样性是北京市京津风沙源低效林改造工程亟待解决的问题。基于此,本研究以北京山区侧柏林为研究对象,基于36个标准样地的实测数据,采用结构方程模型阐述了林分结构因子和环境因子对林下草本物种多样性的耦合影响,以期为北京山区林地森林质量的提升提供理论依据和技术支撑。
1 材料与方法
1.1 研究区概况
研究区为北京市西部的京津风沙源治理工程区,包括房山、门头沟、昌平、延庆,地理位置是:115°23′~115°34′ E,39°15′~40°55′ N。该区域属于温带大陆性季风气候,年均降水量为490~630 mm,年均气温9~13℃。山区地貌类型复杂多样,平均海拔200~450 m,土壤类型以山地褐土和沙壤土为主,土层薄,部分区域土壤肥力较低。工程造林主要以侧柏、油松(Pinustabulaeformis)为优势树种,黄栌(Cotinuscoggygria)、山杏(Armeniacasibirica)、元宝枫(Acertruncatum)等也有少量分布,林下草本植物主要有白莲蒿(Artemisiastechmanniana)、小红菊(Chrysanthemumchanetii)、细叶薹草(Carexrigescens)、中华卷柏(Selaginellasinensis)等。
1.2 样地选取
根据森林二类调查资料,依据侧柏林位置分布的均匀性和代表性,在研究区内选择人为干扰较轻的侧柏林为研究对象,最终选取了36块不同海拔、坡度和土壤的20 m×20 m的标准侧柏林样地。统计整理后立地因子见表1。
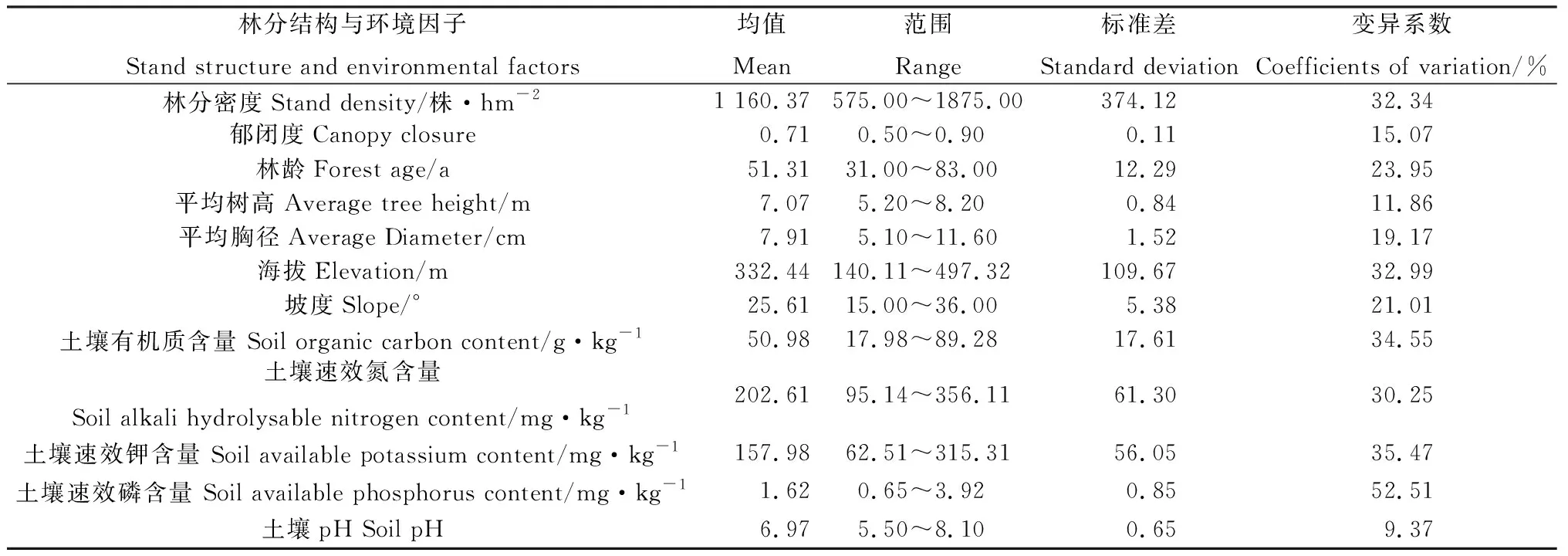
表1 标准样地的林分结构和环境因子数据Table 1 Stand structure and environmental factors data of sample plots
1.3 林分结构及草本植物观测
2021年6月,测量标准样地内所有乔木树种的树高、胸径、郁闭度和林分密度等基本林分结构参数,其中,林分密度通过计算单位面积的林木数量获得,乔木层郁闭度采用样线法调查,按对角线总长与树冠冠幅总长之比计算郁闭度大小。同时,在每个标准样地内随机布设5个1 m×1 m草本样方,并记录各样方内草本植物的名称以及每个物种的株数,用于计算其林下草本植物的多样性。统计整理后标准样地的林分结构因子见表1。
1.4 土壤取样及测试
在各标准样地内随机选择3个具有代表性的土壤取样点,采用100 cm3的标准环刀对(0~10 cm,10~20 cm)土层进行采样,每层土样取3个重复,将野外采集的土壤样品带回实验室测定其有机质、速效氮、速效钾、速效磷含量以及pH值,其中,土壤有机质采用稀释热-重铬酸钾氧化法测定;速效氮采用氯化钾溶液浸提后用流动分析仪测定;速效钾使用火焰光度计测量;速效磷采用碳酸氢钠加活性炭浸润后用流动分析仪测定;pH采用电位法测定,测定结果见表1。
1.5 研究方法
1.5.1多样性指数计算 本研究选用Simpson多样性指数、Pielou均匀度指数、Margalef丰富度指数来表示林下草本植物多样性水平,计算公式如下:
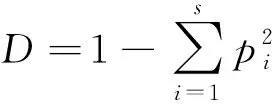
(1)
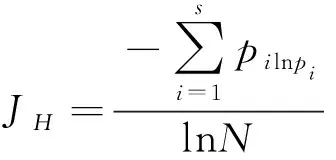
(2)
Margalef丰富度指数H=(S-1)/lnN
(3)
式中,S表示林下草本植物种的数量,pi为第i种个体数占所有种个体总数的比列,N为林下草本植物个体的总数。
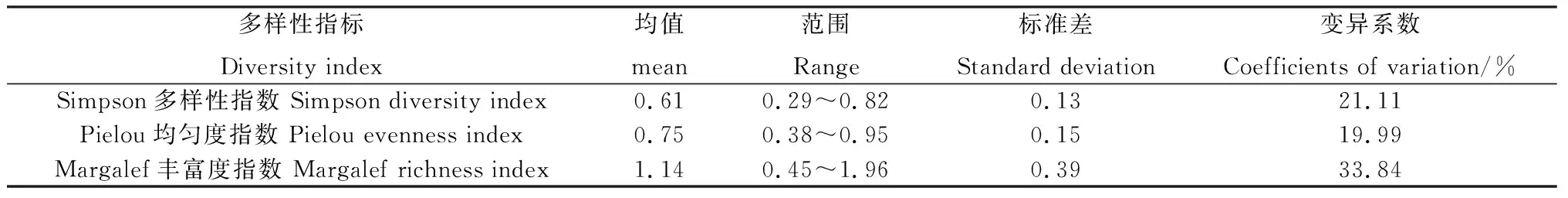
表2 标准样地的多样性指标计算结果Table 2 Calculation results of diversity index of standard sample plots
1.5.2多元线性逐步回归 将多样性指数作为因变量,将林分结构和环境因子作为自变量进行多元线性逐步回归分析,筛选出影响林下草本植物多样性的主导因子。
1.5.3结构方程模型的构建 采用结构方程模型模拟和验证因子间的交互关系以及各林分结构因子和环境因子与林下草本多样性的因果关系。本研究在构建结构方程模型前,采用SPSS软件中的皮尔逊相关系数法(Pearson correlation coefficient)检验各林分结构因子和环境因子与林下草本物种多样性的相关关系,筛选出与林下草本物种多样性具有显著性相关的因子(P<0.05),作为结构方程模型构建的基本变量。模型的精度通过模型的拟合度指标反映,当CMIN/DF值介于1到3之间,RMSEA值小于0.08,CFI大于0.9,显著性概率值P>0.05时表示假设模型的拟合度较高[30]。
1.6 数据处理与分析
本研究使用Excel2018和SPSS Statistics 20进行数据整理、统计、分析和绘图,利用Pearson相关系数法对环境指标和林分结构指标与林下植被物种多样性指标的相关性进行分析,采用Origin 2018进行绘图,应用Amos 21.0软件进行结构方程模型构建。
2 结果与分析
2.1 林分结构因子、环境因子与林下草本植物多样性的相关性分析
由表3可知,林分密度、郁闭度、林龄、海拔、土壤有机质、土壤速效氮与林下草本物种多样性指数具有显著性相关关系(P<0.05),而平均树高、平均胸径、坡度、土壤速效钾含量、土壤速效磷含量、土壤pH值与林下草本物种多样性指数无显著相关关系。其中,表征林分结构的林分密度、郁闭度、林龄与林下物种多样性呈显著负相关,而表征环境的海拔、土壤有机质含量及土壤速效氮含量与物种多样性呈显著正相关(表3)。因此,选择海拔、土壤有机质、土壤速效氮作为环境因子,并将林分密度、林龄、林分郁闭度作为林分结构因子逐个加入结构方程模型。
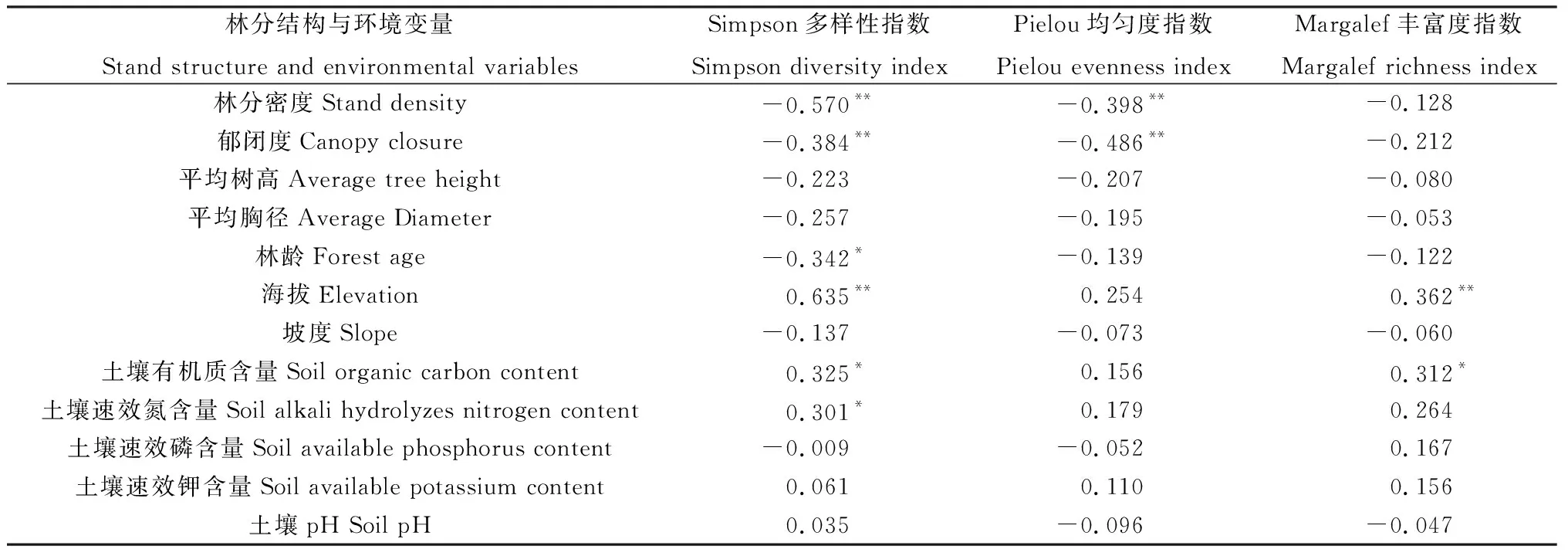
表3 林分结构因子和环境因子与多样性指数的相关性Table 3 Correlation between stand structure factors,environmental factors and diversity index
2.2 林分结构因子、环境因子与林下草本物种多样性的逐步回归分析
由表4可知,各回归方程的拟合系数良好,可用以检验各因子对草本多样性的综合影响,影响林下草本Simpson多样性指数的主导因子为海拔、林分密度、有机质,影响Pielou均匀度指数的主导因子为郁闭度;而影响林下Margalef丰富度指数的主导因子为海拔。

表4 林分结构因子、环境因子与林下草本物种多样性的多元回归方程Table 4 Multiple regression equation between stand structure factors,environmental factors and understory herbaceous species diversity
2.3 林分结构因子、环境因子与林下草本物种多样性的结构方程模型
最优结构方程模型显示,显著性概率P值为0.15、卡方和自由度的比值CMIN/DF为1.32、比较拟合指数CFI为0.96、误差均方根RSMEA为0.07,说明该模型优配度良好,可用于检验林分结构因子和环境因子对林下草本物种多样性的耦合影响。海拔、林分结构和土壤养分对林下草本物种多样性均有影响,总路径系数分别为0.710(P<0.01)、-0.505(P<0.01)和0.311(P<0.05)。海拔与林分结构和土壤的路径系数分别为-0.26和0.32(P<0.05),林分结构与土壤的路径系数为0.15(图1)。
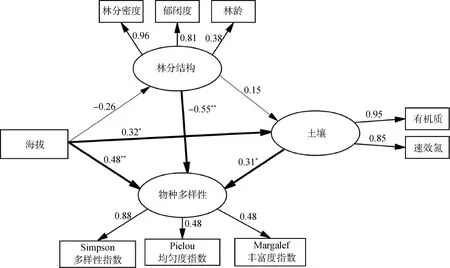
图1 林分结构和环境因子对林下草本物种多样性影响的结构方程模型Fig.1 Structural equation model of effects of stand structure and environmental factors on understory herbaceous species diversity注:利用粗线和细线箭头显示各变量间因果关系的显著性,粗线箭头表明两个变量间的因果关系具有显著性,细线箭头表明不具备显著性,* P<0.05,** P<0.01Notes:Use thick and thin arrows to show the significance of causality among variables. Among them,the bold arrow indicates that the causal relationship between the two variables is significant;The thin line indicates that it is not significant,* P<0.05,** P<0.01
海拔和林分结构对林下草本物种多样性不仅存在直接影响,还通过影响林分结构和土壤养分对林下草本物种多样性产生间接影响。海拔和林分结构对林下草本物种多样性的直接效应为0.478和-0.550,间接效应为0.232和0.046 (表5)。结构方程模型分析结果表明林分结构和海拔对林下草本植物多样性的影响起决定作用(P<0.01),土壤养分对林下草本植物多样性起重要作用(P<0.05)。

表5 林分结构因子、环境因子对林下草本物种多样性的标准化影响效应Table 5 Standardized effects of stand structure factors and environmental factors on understory herbaceous species diversity
3 讨论
林下草本植物多样性的维持机制始终是生态学研究的核心和热点问题之一,明确影响林下草本植物多样性的主导因素对森林生态系统的可持续经营具有重要意义[31-32]。结构方程模型(SEM)对于验证多因素耦合影响过程具有巨大优势,目前已被广泛应用于环境和生态学研究中。We等[27]采用结构方程模型研究了区域地形和林分结构对土壤特征的影响;Hou等[29]利用结构方程模型研究了降雨期间控制水土流失的最佳植被覆盖和种植坡度,其研究结果充分体现了模型的科学性和可靠性。本研究将林分密度、郁闭度和林龄表征为林分结构因子,将海拔、土壤有机质和土壤速效氮表征为环境因子,采用结构方程模型探讨了林分结构因子和环境因子对林下植物多样性的直接和间接影响,模型结果显示,表征环境因子的海拔对林下草本物种多样性具有极显著影响(P<0.01),且同时存在直接效应和间接效应,但直接效应强于间接影响效应,这与王德君等[33]研究结论基本一致,这可能是因为海拔通过调节林下草本物种生长发育所必须的温度、光照、水分而直接影响林下草本的物种多样性,通过改变土壤养分及土壤微生物变化间接影响了林下草本的生长和发育[34]。相关研究也表明,海拔梯度的变化包含了水、热、光照等多环境因子的综合变化,这些变化直接或间接地改变了林下土壤理化性质和植物的生长发育[35-37]。低海拔干旱限制了草本的生长和繁殖,只有中海拔地区具有适当的温度和降水组合,才促进了林下草本物种多样性[38]。
土壤作为植物生长必不可少的条件之一,其养分含量的高低直接决定着林下草本植物的组成和分布[20]。本研究发现,土壤养分因子对林下草本物种多样性的影响显著(P<0.05),仅存在直接影响效应,其中,表征土壤因子的有机质和速效氮对林下草本物种多样性的影响相对较高,这与胡冬等[14]认为林下草本植物多样性主要受土壤有机质和速效氮的影响结论基本一致。这可能是因为土壤有机质作为草本植物主要的养分来源,而氮作为草本植物生长所必须的大量营养元素,在促进草本植物生长、群落演替、生态系统结构稳定方面具有重要作用[39]。但部分学者[40]认为土壤有机质和速效氮与林下草本植物多样性无显著关系,这可能是研究样地的立地类型和树种存在差异。
本研究还发现,林分结构因子对林下草本物种多样性的影响极显著(P<0.01),且同时存在直接影响和间接影响,但直接影响强于间接影响。其中郁闭度和林分密度的作用相对较大,而林龄的作用相对较小。Wagner等[41]研究认为,乔木层的郁闭度是制约林下草本物种多样性的主要林分结构因子,与本研究的结论基本一致,这可能是因为乔木层的郁闭度直接决定林内的有效光强度,对林内光、热、水、肥等资源具有再分配作用,从而直接影响林下草本物种的分布和个体生长,特别是对一些喜阳和非耐阴植物,随着光照强度的变化,其种群发育会发生改变,进而改变了林下草本的物种多样性[42]。另一方面,在低郁闭度的林内,光照强度的增加促进了林下枯落物的分解和土壤微生物活动,加快了土壤有机质的形成,进而有利于草本植物吸收更多的土壤养分[43-44],从而间接的改变了林下草本物种的组成和分布。林分密度也是制约林下草本植物多样性的主导因子(P<0.01),且与林下草本植物多样性呈负相关关系,这与张柳桦等[25]认为林分密度是决定林下植物多样性的主导因子,且随着林分密度的增加,林下草本植物多样性逐渐降低研究结论基本一致,这可能是因为林分密度直接决定着林内光、热、温度、湿度、土壤养分等一系列草本生长环境[45-47]。一方面,高密度的侧柏优势树种抑制了林下草本对光的吸收效率,使得林下喜阳草本植物逐渐消失,从而直接影响林下草本物种多样性,另一方面,高密度的优势树种充分吸收了土壤养分,加剧了对林下草本物种的生态位竞争,间接的影响了处于劣势的草本植物生长[46]。但有学者[41]认为,林下草本物种多样性与林分密度无显著关系,与本研究的结论产生一定的差异,这可能是研究样地的立地条件、树种组成、林分密度等因素存在差异。因此,今后的研究应注重考虑林分结构和立地条件对林下物种多样性的耦合影响。本研究发现林龄对林下草本物种多样性的影响相对较小,但许多学者[48-50]认为,林龄也是制约林下草本物种多样性的主导因素,与本研究的结论产生一定的差异,这可能是本研究的侧柏林基本接近中、成熟林,对于幼林龄侧柏林下草本物种多样性尚未研究,因此,在今后的研究中,可注重研究整个侧柏生长周期的林下物种多样性变化,为森林生态系统的协调发展提供更多的科学理论依据。
合理的林分结构是提高森林生态系统服务功能的关键,而北京山区地形地貌条件较为恶劣,如何提高侧柏林下物种多样性对低效林改造工程具有重要意义。本研究发现林分郁闭度和林分密度是影响林下草本植物多样性相对较高的林分结构因子。因此,欲维持林下草本物种多样性,可将郁闭度和林分密度调整为0.65~0.71和1 000~1 125 株·hm-2,林下草本物种多样性整体水平较为稳定,适当补植阔叶树种提高其混交度,形成结构复杂的复层针阔混交林。本研究在利用结构方程模型时未充分考虑气温、降水等气候变量是否会对侧柏林下草本植物多样性产生影响,因此,在今后的研究中,应考虑获取更多的环境变量数据,且增加样本的数量,全面系统地解析影响侧柏林下草本植物多样性的维持机制。
4 结论
海拔与林分结构是影响北京山区侧柏林下草本植物多样性整体水平的主导因素,且直接影响为主要作用。土壤对林下草本物种多样性整体水平具有重要影响,但不是主要影响因子,仅存在直接影响。影响多样性指数的主导因子因各指数差异而异,其中,影响林下草本Simpson多样性的主导因子为林分密度、土壤有机质、海拔,影响林下草本Pielou均匀度指数的主导因子为郁闭度,影响林下草本Margalef丰富度指数的主导因子海拔。因此,欲维持和提高林下草本物种多样性整体水平,可适当调整林分密度和郁闭度,维持合理林分结构,并适当的提高土壤养分。
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
少儿美术(2020年8期)2020-11-06 05:21:38
防护林科技(2020年6期)2020-08-12 13:34:40
福建林业(2020年5期)2020-03-18 08:23:02
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
热带林业(2019年4期)2019-03-05 09:53:58
森林工程(2018年3期)2018-06-26 03:40:46
亚热带资源与环境学报(2015年3期)2015-01-22 07:05:36
