
家禽产肉性能相关基因研究进展
2022-08-25 09:25向炜萱彭佳诚胡志刚刘小林
家畜生态学报 2022年7期
向炜萱,彭佳诚,胡志刚,刘小林
(西北农林科技大学 动物科技学院,陕西 杨凌 712100)
禽肉作为人类摄取动物蛋白的理想食品来源,其产肉性能得到了人们极高的关注。家禽的肉用性状呈不连续性变异,是受微效多基因控制的数量性状,主要包括体重与体增重、屠体性能、体型和骨骼指标(肉用家禽主要是胸宽和胸角度)等。家禽不同生长、发育阶段的肌肉受到了复杂的细胞外界多因子和细胞外基质调控。研究不同阶段家禽生长发育过程中产肉相关性状基因的作用机理,对提高我国肉用禽类生产能力有着关键的作用。近年来对产肉性能相关基因的研究包括肌肉生长抑制素、生肌调节因子、生肌细胞增强子因子家族2和基因家族(3、7)等。本文通过概述近年来肌肉发育相关基因调节机制的研究,为提高禽类肉品质提供关键候选标记基因,为我国地方家禽品种改良和选育提供参考。
1 家禽生长性能与肉用性状
我国地方畜禽品种资源丰富、肉质较为鲜美,但产肉性能表现差异较大。比如全净膛率,浦东鸡为73.90%,黑羽仙居鸡为66.65%;大围山微型鸡和武定鸡的胸肌率达18.76%,而略阳乌鸡胸肌率仅为16.91%;在鸭上,巢湖麻鸭(公)的屠宰率可达88.07%,高邮鸭为85.34%。其它生长性能见表1。另一方面,和一些引进的品种相比,我国地方家禽品种呈现出生长速度更慢、体格更加瘦小等问题。例如,与生长速度快、肉质优良的白羽肉鸡相比,我国藏鸡体型较小、生产性能不高;樱桃谷鸭、番鸭等大型肉鸭体重显著高于肉蛋兼用性地方品种(花边鸭和四川麻鸭),且肉品质以及屠宰性能也显著高于地方品种。
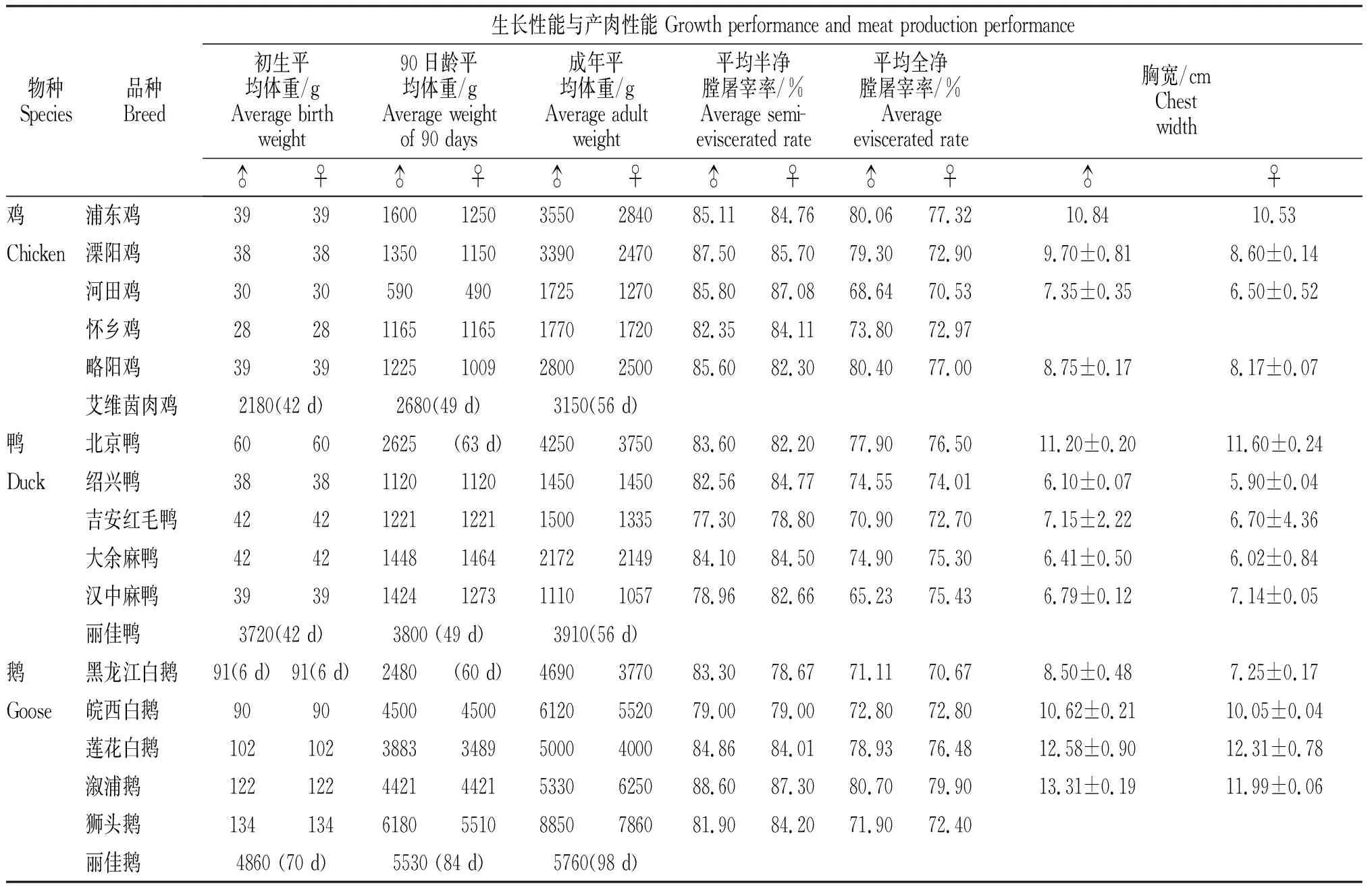
表1 中国地方家禽品种以及外来家禽品种生长性能与产肉性能统计[16]Table 1 The statistics on growth performance and meat production performance of Chinese indigenous and foreign poultry breeds
2 家禽肌肉发育过程
家禽的肌肉主要分为心肌、平滑肌以及骨骼肌,均来自于中胚层组织。从胚胎发育开始,体节细胞会经过细胞增殖、分化等步骤,成为多核肌管,最终发展成肌肉组织。体节细胞在收到胚胎生长发育信号后被诱导分化为成肌细胞,成肌细胞不断分裂增殖,迁移到胚胎不同部位继续增长,黏附并融合形成多核肌管,继续分化形成初级纤维,再在初级纤维形成的位置上分化为次级纤维。最后,次级纤维通过肌节的收缩单位“Z”带连接为成熟肌纤维。目前,与家禽孵化期肌细胞增生有关的调节信号途径、出生后肌纤维增生和肥大的细胞外因子调控机制以及孵化期和孵化后肌肉增长的关系还有待进一步研究。动物个体在出生时,就几乎完成了全肌纤维的发育,且实际的肌纤维总量也都是确定的。在出生之后,动物骨骼肌生长依靠合成大量的蛋白质使得肌纤维肥大。进入到肌纤维中的卫星细胞,会在肌肉生长的整个过程中发挥重要作用。肌纤维的生长、肥大与多种因子如胰岛素样生长因子、肝细胞生长因子、β-转化生长因子等有关,同时也与细胞外基质有一定关系。
3 家禽肉用性状相关基因的研究
3.1 肌肉生长抑制素(MSTN)
肌肉生成抑制素(myostatin,)是转化生长因子β-超家族中的一种因子。在肌肉发生、分化生长、萎缩过程中均起作用,其表达会抑制肌肉生长,控制肌肉发育处于正常水平。调节该基因实际表达量能够控制肌肉细胞面积,让肌肉纤维组成、重量都受到直接或间接的影响。Liu通过比较云南武定鸡和艾维昂肉鸡中的表达量,揭示了生长更快速的肉鸡品种中mRNA表达量显著低于武定鸡。梁耀伟在鸡、鹌鹑及其杂交后代的研究中提到,通过阻止成肌细胞从G1到S期的细胞循环来抑制肌肉发育,且mRNA表达量随成肌细胞生长周期循环而变化。
目前关于在家禽上的研究侧重于育种工作,经常将其作为家禽肌肉性状选择的关键标记基因。廖娟等通过测量黄杂鸡的体尺、体重以及检测基因启动子区SNP,发现了基因启动子区一个SNP位点: c.-1782 A>T与体重、体斜长有着密切关联,可将该SNP作为黄杂鸡育种工作的分子遗传标记之一。王彦钦等通过测量拜城油鸡的生产性能和基因多态性,证明了基因第1外显子c.234G>A突变显著地影响12周、18周、24周实验鸡的宰前活重、屠体重、胸肌重、腿肌重、半净膛重和全净膛重。Ye等在三种优良商品肉鸡上证实了中的非同义突变 (g.4842T>G)和体重有关。由此可见,基因突变失活往往使得家禽个体肌肉率上升,肉质提高。
Bhattacharya等表明在鸡的发育过程中的敲除会导致肌肉发育不正常、体重显著增加,且CRISPR技术被证实可有效的用于活鸡肌肉组织基因编辑。杨永平对基因进行了测序、序列分析,提出至少有7种类型的剪接体存在于鸭基因当中,其中剪接因子U2AF35可能对骨骼肌卫星细胞存在调控作用从而影响肌肉生长性状。这从基因编辑的角度提供了选育方法。
胰岛素生长因子Ⅰ (-Ⅰ,-Ⅰ)和基因具体表达水平与家禽肌细胞生长周期有关。汤青萍表示促进肌肉发育的-Ⅰ和抑制肌肉发育的MSTN蛋白表达水平同时上调和下降。胡艳通过测定高邮鸭和金定鸭2个不同品种胸腿肌的发育情况与-mRNA表达的关系,发现骨骼肌生长速率与-Ⅰ-mRNA 表达比值的变化曲线吻合,肌细胞增殖则比值降低,肌细胞肥大则比值升高。姬改革发现,高邮鸭和金定鸭-Ⅰ与mRNA 的比值有显著差异,均呈显著负相关,这样的差异极大可能会导致品种间鸭胚胸肌发育程度不同。可以得出结论,家禽肌肉发育尤其是骨骼肌生长速率受到与-Ⅰ协同控制,推测这两个基因在筛选肉用性状中起重要作用。
3.2 生肌调节因子(MRFs)
生肌调节因子(MRFs)包涵了4种肌肉特异性调节因子,即Myf5、MyoD、MRF4(别称Myf6)、MyoG,他们的表达在肌肉生长、发育的完整过程中起着直接激活细胞向成肌细胞转化的作用。基因家族都有一个保守的-结构,是决定其重要生物学功能的调控区域。Yin等在2015年发现,运动使得基因家族4个因子的mRNA丰度增高,从而导致肌肉纤维直径增加,最终影响成年鸡骨骼肌的肉质。
5基因是肌肉生长过程中最早表达的,该基因会激活肌前体细胞多项分化功能。张涛在京海黄鸡上发现,出生后5表达量随鸡的年龄增长呈下降趋势,这说明5主要在家禽胚胎和早期肌肉发育中起作用。同时5被证实在生长性状优异的个体上表达量也高。
基因的表达,会让M-肌酸激酶、肌原蛋白I、肌球蛋白等多个基因都被激活。张久盘等发现,表达峰值出现在固原鸡体成熟前。因和5作用时间段重叠,且通过Rudnicki的研究发现,和5作用相互补充并在同一条基因通路上游启动骨骼肌生成,所以往往将二者联合研究:Zhang等研究表明,藏公鸡的胸肌、大腿肌中、5的mRNA表达量与体长、胸宽、胸深、胸骨长、胫长、胫围和嵴宽等生长性状呈正相关。陶志云等对高邮鸭、金定鸭的研究表明,5、1表达量虽然存在品种和发育时期表达量差异,但高峰均在17胚龄。以上试验均证实了5、可以决定成肌前体细胞的命运和促进肌肉细胞增殖。
4在肌管生长、成熟时期发挥着重大作用,基因主要在成肌细胞分化、融合成肌纤维的过程中发挥作用,而二者被证实共同作用于肌肉分化。张久盘等在固原鸡上得出结论,肌肉中表达量极显著高于其他所有组织,其中胸肌表达量高于腿肌,但同时在10日龄表达量达到丰度,证明促进早期细胞分化,激活肌肉细胞生长,促进肌肉发育。李丰耘等研究发现,的相对表达量与盐津乌骨鸡和大围山微型鸡的屠宰性状、生长性状显著相关,且在体型相对较大的盐津乌骨鸡表达量更高,由此可将作为研究地方禽种体型大小的关键基因。
3.3 生肌细胞增强子因子家族(MEF2)
脊椎动物中,肌细胞增强因子-2家族主要包含下述四个成员:2、2、2、2。2广泛存在于骨骼肌、心肌、平滑肌中,2可结合DNA序列来影响生长因子,并具有组织特异性,在骨骼肌和心肌细胞中表达。2被证实其通过改变蛋白质分子构象来参与骨骼肌、心肌基因转录。2在肌管形成期有大量的表达,而多层肌肉母细胞2表达量极高,因此都被认为与骨骼肌的分化生长有关。
易恒洁发现,三穗鸭2基因的g.47915G>A、g.47918G>A两种不同基因型会显著(<0.05)影响全净膛率,即基因型全净膛率显著高于基因型(<0.05)。这和赵忠海对于这两个基因SNP位点的研究结果一致。除此以外,第11外显子的g.47915G>A、g.47918G>A 位点,基因型会对全净膛率产生正向加性效应。周艳提出,肉鸡活体质量、屠体质量等和肉质性状有关的指标,都与2基因的5′非翻译区、第4外显子、第7内含子内的3个SNPs位点有明显联系。师新彩等指出,对于兴义鸭的活重、屠体重、净膛率、胸肌重、腿肌率等多个产肉性能指标而言,2基因有4个SNP位点(16236、16305、16359、21071)会带来极大的影响。由此2家族已被作为影响鸡肉质性状的候选基因。
3.4 Pax基因家族(Pax3、Pax7)
基因家族编码的蛋白质是重要的转录调控因子,其结构特点有着由128个氨基酸构成的保守成对域结构域。基因家族进一步可分类为:1、9;2、5、8;3、7;4、6四个亚家族。3被证实调控脊椎动物胚胎期的四肢发育,而7在肌肉损伤的时候起显著作用,二者在肌肉发育过程中都作用于卫星细胞,促进卫星细胞的生长、融合,从而使肌纤维肥大,促进动物肌肉的发育。
常国斌通过统计3基因在如皋鸡上不同生长时期的表达量发现,该基因在胚胎期的表达量显著高于出生后,且存在一定组织特异性。王吟方亭研究了浙东白鹅3和7基因表达规律,发现3基因在胚龄生长阶段表达量最高,随着肌肉生长发育表达量逐渐降低,且7在肌祖细胞进入骨骼肌时期发挥关键作用。张荣萍以黑鹜鸭与北京鸭为研究对象,从分子水平证实了3、7表达量存在发育时期差异性和组织特异性。以上结果可使今后研究中针对性提高不同时期和不同组织的低基因家族表达量。张锁在固始鸡安卡鸡F2代里筛选出了两个7的SNP位点,证实其与鸡的屠体等肉质性状有关,为鸡7提供了重要的变异位点。
4 小 结
肌肉生长因子中的、、2、基因家族(3、7)的位点和不同组织、不同时期的表达量对我国地方家禽体重、屠宰率等性状有影响,以上基因的某些多态位点可作为提高家禽产肉性能的分子标记,且生肌因子的表达量高的组织或时期,往往肌肉性能更优秀;抑制肌肉发育的因子表达量低或位点失活后,往往也能提高肉质和肉产量。可以运用高通量测序技术、转录组分析技术并结合生物信息学方法筛选、检验更多的SNP位点。随着禽类越来越多产肉相关基因位点被发现,在今后的研究中应将这些基因之间的互作作为工作重点,并确认其在肌肉发育中生效的具体时期,有针对性提高或降低基因表达量,为家禽品种选育提供分子机制的研究思路,实现中国地方家禽品种肉质改良,提高现代化家禽养殖业的经济效益。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
中国农业科学(2022年15期)2022-08-09
电脑报(2020年40期)2020-11-06
电脑知识与技术(2018年19期)2018-11-01
农民致富之友(2018年3期)2018-03-28
农民致富之友(2018年3期)2018-03-28
农民致富之友(2017年14期)2018-01-27
现代仪器与医疗(2017年1期)2017-03-29
山东体育学院学报(2016年2期)2016-05-25
