
植物叶功能性状对光伏组件遮阴的响应与生态策略*
2022-08-23 02:46罗久富罗忠新赵世麒吴赛男罗小林隋欣
西部林业科学 2022年4期
罗久富,罗忠新,赵世麒,吴赛男,罗小林,隋欣
(1.中国水利水电科学研究院 ,北京100038;2.国家水电可持续发展研究中心,北京100038;3.国家电投云南国际电力投资有限公司,云南 昆明 650228)
云南省70%以上的县年太阳能辐射总量高于5 000 MJ/m2[1],光伏发电潜力巨大。但是,在青藏高原隆起、东南和西南季风共同作用,以及人类活动的共同影响下,在云南省内形成碳酸盐岩出露严重、岩溶发育强烈、生态系统极为脆弱的石漠化区[2]。将石漠化区植被恢复与光伏发电结合,推广“新能源+生态修复”模式[3],是发展生态光伏的重要环节。
光伏板下太阳辐射减少约25%~95%[4],长波入射升高、天空视域因子降低驱动土壤中能量流动异质性变化[5]。板下区域土壤水势和水分消耗速度降低、土壤可利用水分含量增加[5]。土壤温度时空变异规律发生改变,在季节尺度上变异系数降低[6],削弱了夏季高温或冬季低温对植物生长的胁迫。伴随着能量流动和物质循环,土壤粗糙度增加[7]。
生境异质性影响植物功能性状构建和生存策略。叶片作为同化作用关键载体和响应环境资源变化最敏感的器官[8],沟通地上与地下物质交换和能量流动。叶表型可塑性驱动植物应对异质性生境采取积极或保守等不同的生态策略,植物通过调节叶面积、厚度、比叶面积等功能性状应对不同的资源环境,平衡其生存、生长与繁殖过程[9]。例如,弱光环境驱动植物叶片变大变薄来获取更多的光资源[10],叶变小增厚抵抗强光胁迫[11],也有研究指出光资源变化对植物的叶厚等功能性状无显著影响[12]。在生态脆弱地区的植被恢复初期,植物的生态策略影响群落演替方向。但目前关于光伏对植物的影响多集中在板下或温室农作物的产量及品质方面[13],针对生态脆弱区光伏场区植被恢复的研究处于初步阶段[14-15]。光伏板和石漠化双重影响下的植物适应性机制尚不明晰,限制“双碳”目标下生态光伏发展。
因此,本研究基于植物叶功能性状反映其生态策略的理论基础[16],探索石漠化区光伏电站场区异质性生境下植被恢复物种的生态策略。本研究选用乡土能源植物和花境植物对光伏电站场区进行植被恢复,监测光伏板间和板下的植物叶片功能性状,探讨植物在异质性生境中的生态策略,以期拓宽石漠化光伏场区植被恢复的物种选择范围,促进区域地表状况稳定、植被恢复和区域生态功能提升,为创新传统的石漠化脆弱区植被恢复与能源供给结构、推广“新能源+生态修复”模式提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于云南省昆明市东川区“生态修复+光伏”示范基地(103°8′30.56″E、26°9′55.70″N),海拔约1 280 m,基地所在东川区年均温14.9 ℃,年均降水量约1 000.5 mm,集中在5-9月,年蒸发量1 856.4 mm。年日照2 327.5 h,属于太阳能较丰富区。场区土壤类型以红壤为主,植被稀疏,以草本植物为主,植物群落学调查显示:扭黄茅(Heteropogoncontortus)为建群种,伴生狗肝菜(Diclipterachinensis)、猩猩草(Euphorbiacyathophora)、截叶铁扫帚(Lespedezacuneata)、假杜鹃(Barleriacristata)、戟叶酸模(Rumexhastatus)等。
1.2 试验设计及样品采集分析
2020年完成光伏电站建设并投入运营,光伏板前檐距地面垂向高度2.5 m,具备小灌木生长空间条件。筛选具有植被修复、固碳释氧、固土保水等功能的能源植物与花境植物鹅掌柴(Scheffleraheptaphylla)、红花檵木(Loropetalumchinensevar.rubrum)、假连翘(Durantaerecta)和黄金菊(Euryopspectinatus)对场区进行植被恢复[17-18],各植物选取长势分别一致、生长良好的营养袋苗,通过集水灌溉技术进行管护。
2021年10月,在光伏板间和板下中央布设样品采集区,对4种植物进行样品采集。在各采集区随机选择15株无病虫害、长势均匀的个体,共120株个体,采集冠层当年生枝条3~5枝,装入自封袋,迅速带回实验室测量指示植物资源利用、抗逆性等功能的相关性状。
每株植物贴枝条表皮剪取枝条中部成熟、完整、健康的叶片1片,5片为1组称量叶鲜质量(leaf fresh weight,LFW,g),鹅掌柴的掌状复叶剪取中央小叶测量。测量叶厚(leaf thickness,LT,mm),避开叶脉选取叶上、中、下部位测量取均值为叶厚。叶厚测量完成后利用EPSON V39扫描仪(Indonesia)扫描叶片,保存像素300 dpi,随后于105 ℃条件下杀青30 min后,75 ℃条件下烘干至恒重,称量干质量(leaf dry weight,LDW,g),计算叶含水率(leaf water content,LWC,%)。利用Photoshop CC 分析叶面积(leaf area,LA,cm2),计算比叶面积(specific leaf area,SLA,cm2/g)、叶体积(leaf volume,LV,cm3)、叶组织密度(leaf tissue density,LTD,g/cm)。表型可塑性指数(phenotypic plasticity index,PPI)采用Ashton所提的计算方法[19]。每种植物重复3次。相关计算公式如下。
式中:PPI(A)表示某个性状A的可塑性指数,0≤PPI(A)≤1;Amax和Amin表示性状A的最大值和最小值。
1.3 数据处理与分析
利用Microsoft Excel 2016对原始数据进行预处理,利用one-way ANOVA检验不同指标的差异性,利用two-way ANOVA 分析植物种类差异和采集区环境差异对植物叶功能性状的影响,数据分析在SPSS 25.0中实现;利用Origin 2016对方差分析结果进行图形化展示。
2 结果与分析
2.1 物种和生境差异对叶功能性状的影响及其可塑性
从表1可见,物种和生境差异对叶功能性状影响显著。4种植物的叶功能性状在物种间差异显著,各功能性状在异质性生境中也差异显著,但是物种和生境的综合效应对叶含水率(P=0.308)、叶组织密度影响不明显(P=0.607)。
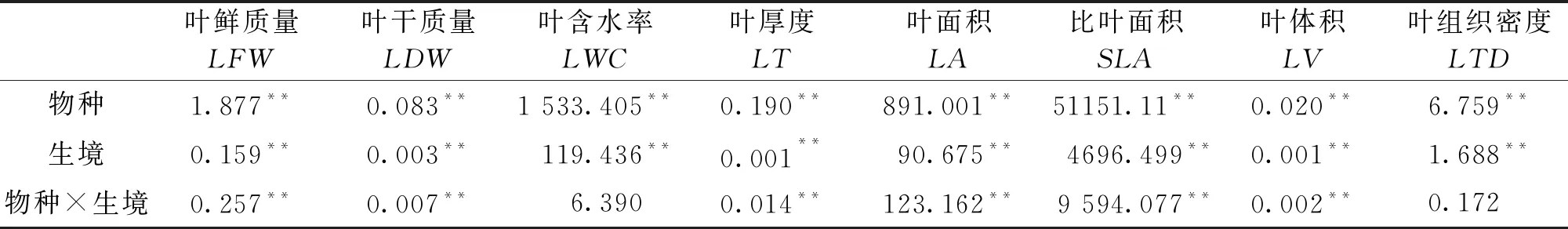
表1 光伏阵列不同光照区植物叶功能性状的双因素方差分析(F值)
8个叶功能性状平均可塑性指数大小为黄金菊>鹅掌柴>假连翘>红花檵木(表2)。红花檵木可塑性指数远低于其余3种植物,惰性较强,惰性响应性状数量最多。叶鲜质量(LWC)为4种植物共同的惰性响应性状(PPI<0.20),黄金菊、鹅掌柴、假连翘可塑性指数最小的均为LWC,红花檵木可塑性指数最小的是LT。黄金菊、鹅掌柴、假连翘、红花檵木最大可塑性指数分别是LV、LV、LDW、SLA,对环境变化具有最敏感响应性。

表2 植物叶功能性状的表型可塑性分析
2.2 光伏板遮阴对叶水分储存及利用功能性状的影响
光伏板遮阴后,鹅掌柴和黄金菊LFW、LDW显著增加(P<0.05),假连翘LFW、LDW降低(P<0.05),红花檵木LFW、LDW变化不明显(图1 A~B);4种植物的LWC增加,其中鹅掌柴、红花檵木、黄金菊LWC增加显著(图1 C);鹅掌柴和黄金菊LT增加,而红花檵木和假连翘LT降低,且黄金菊和假连翘LT变异显著(图1 D)。
各物种对光伏板遮阴的响应差异改变物种间叶水分存储和利用的差异性。光伏板遮阴后,黄金菊-红花檵木LFW、LDW、LT,黄金菊-鹅掌柴LWC由不显著转变为显著差异(图1 A~D)。

图1 光伏阵列不同光照区植物叶功能性状差异(平均值±标准差)
2.3 光伏板遮阴对叶光、碳资源获取功能性状的影响
光伏板遮阴增强植物叶片资源捕获功能。光伏板遮阴后,4种植物的叶面积(LA)均呈增加趋势,且鹅掌柴和黄金菊分别增加79.47%和82.66%(P<0.05)(图1 E);鹅掌柴、红花檵木和假连翘的SLA均呈增加趋势,其中假连翘增幅45.50%(P<0.05),黄金菊的比叶面积(SLA)呈微弱的降低趋势(P>0.05)(图1 F)。LA和SLA的种间异速变化改变物种间的性状差异,红花檵木-假连翘和红花檵木-黄金菊LA差异、鹅掌柴-红花檵木和假连翘-黄金菊SLA差异由不显著转变为显著,而红花檵木-黄金菊SLA差异性降低(图1 E~F)。
2.4 光伏板遮阴对叶综合投资及防御功能性状的影响
光伏板遮阴改变植物种间叶综合投资差异,降低了叶片机械防御功能。光伏板遮阴后,物种间叶体积(LV)响应差异明显,鹅掌柴和黄金菊的LV增幅分别达76.37%和100%,而假连翘LV降低40.12%,红花檵木LV无变化(图1 G);LTD均呈降低趋势,其中黄金菊LTD极显著降低(P<0.01)(图1 H)。
3 讨论与结论
3.1 讨论
3.1.1 石漠化光伏场区植物叶功能性状表型可塑性差异
光伏电站建设和运营过程对环境资源的再分配增强场区生境异质性。叶表型可塑性反映植物对生境的适应策略,是研究植物生态适应性、资源利用策略、物质分配机制等生态策略的关键特征[20]。叶片作为植物获取环境光、热、碳等资源最大的功能器官,通过形态建成变化适应异质性环境资源[10]。本研究参试的4种植物8个叶功能性状表型可塑性综合指数大小为黄金菊>鹅掌柴>假连翘>红花檵木,说明不同的植物受资源需求和获取能力差异,导致叶功能性状可塑性不同[21]。黄金菊除SLA可塑性在4种植物中最低以外,其余性状表型可塑性均最大,而红花檵木叶片表型可塑性处于较低水平。植物种内LWC变幅较小,而LFW、LDW、SLA、LV等具有较高的可塑性,表明叶片LWC与4种植物的适合度均密切相关,此外鹅掌柴和红花檵木的LT、假连翘的LA也与其适合度密切相关[22]。各功能性状的表型可塑性的差异说明植物通过敏感响应性状较高的适应性保障LWC等关键性状的相对稳定,采取在具有不同适合度性状之间进行整合和权衡的“Bradshaw-Sultan”效应生态策略[23]。例如光伏板下黄金菊更容易通过调节植物叶表型性状保障光照、水热等资源获取,提高植物体的生存和繁殖能力,而红花檵木叶功能性状惰性较强,反映其较窄的生态幅[24]。
3.1.2 石漠化光伏场区植物叶功能性状及其生态策略差异
植物表型可塑性驱动植物改变功能性状建成反馈生境变化,形成应对环境的最佳功能性状组合[25]。多数研究指出,弱光环境下植物叶片变薄、干物质含量降低,如三七(Panaxnotoginseng)等在光照降低后植物SLA增加[26]。也有研究指出,弱光会刺激植物LA、LT、LDW增加,SLA降低,如沙棘(Hippophaerhamnoides)叶片对光照强度的降低环境采取叶片变大、变厚的策略[27]。
本研究中在光伏板间强光照环境下,4种植物具有高LTD、低LWC和LA以保障水分固持、提升导热能力并降低蒸腾,增强其应对强光照、干旱环境等非生物因素的防御力和自我保护[28]。光伏板遮阴后,4种植物LWC和LA增加,说明4种植物通过叶片增大来捕获更多的光资源,为其生长提供机会[29-30]。荫蔽环境降低了植物受光照和风等机械胁迫,蒸腾作用降低,在水力学驱动下,植物LTD降低,防御物理损伤的功能下降,叶片周转生长速度增加[31-32]。这种性状组合说明光伏板下植物倾向于资源获取型的生态策略。
但是,植物对资源投资的权衡存在种间差异。鹅掌柴和黄金菊通过增加LT、LA、LFW和LDW的“快速投资-收益”来实现物质积累以应对遮阴环境。黄金菊的SLA降低说明其对物质积累的投入强于叶面积形态构建,植物生长速率增加,叶生物量增加。两种植物形成了叶增大、增厚、养分储存增加的快速资源获取和生长的积极策略[33]。这种策略与本研究中红花檵木和假连翘、前期在库布齐沙漠光伏板下的沙打旺(Astragalusadsurgens)叶片变薄的策略不同[34]。板下红花檵木和假连翘LDW、LFW、LT降低,反映了两种植物的物质积累能力较弱,具有“缓慢投资-收益”的保守型生态策略[33]。叶面积、比叶面积的增加反映其获取光、碳资源的能力提升,但是与鹅掌柴和黄金菊叶面积显著增加不同,红花檵木和假连翘的叶面积增加并不明显,说明其资源获取能力仍然受到限制[35],所以SLA增加可能是建立在牺牲叶片物质积累效率的基础上而形成,植物在叶形态构建和物质积累方面投入均低于板间生境区,其中假连翘受生境变化影响更显著。因此,红花檵木和假连翘应对光伏板遮阴采取了积极和保守协同的生态策略。
本研究中4种植物叶功能性状对光伏板遮阴的不同的响应和适应机制来源于物种、生境特征差异及其交叉影响的综合效应。种间叶功能性状变异的多样化反映了多种生态策略的共存,通过资源保守和资源获取生存策略的稳定组合,提升石漠化脆弱区光伏场区植被恢复效益和生态功能[32,36]。
3.2 结论
石漠化光伏场区植物叶片功能性状及生态策略研究发现,叶表型可塑性黄金菊>鹅掌柴>假连翘>红花檵木。4种植物LFW、LDW、SLA、LV等功能性状具有较高可塑性,LWC是4种植物共有的惰性响应性状。光伏板间植物形成了高LTD、低LWC、LA的性状组合,提高其对板间强光、缺水生境的适应性;光伏板遮阴后,鹅掌柴和黄金菊形成了增加LT、LFW、LWC、LA、LDW和降低LTD等促进物质积累的性状组合,采取资源获取型生态策略;而红花檵木和假连翘形成SLA、LWC、LA增加,LFW、LDW、LT、LTD降低等一系列获取资源但物质积累速率较低的资源获取和保守协同的生态策略。因此,在光伏板下宜筛选具有较高可塑性的资源获取型、板间筛选资源保守策略为主的植物构建群落,提升石漠化光伏场区的生态功能。
猜你喜欢
今日农业(2022年16期)2022-09-22
——以凤凰县为例
湖北林业科技(2022年4期)2022-09-19
河南农业(2022年8期)2022-09-07
——以贵州威宁县为例
测绘地理信息(2022年2期)2022-04-02
绿色科技(2022年4期)2022-03-21
昆明医科大学学报(2022年1期)2022-02-28
今日农业(2021年21期)2022-01-12
今日农业(2021年7期)2021-11-27
大众科学(2017年7期)2017-08-18
建筑建材装饰(2016年14期)2017-01-05
