
RNAi沉默N-cadherin通过MEK-ERK信号通路抑制卵巢癌细胞增殖、侵袭、迁移及上皮-间质转化过程
2022-08-23 02:16陈虹潘琦文李建湘蓝燕潘彩萍
西部医学 2022年8期
陈虹 潘琦文 李建湘 蓝燕 潘彩萍
(河池市人民医院,广西 河池 547000)
卵巢癌是女性最常见和最致命的恶性肿瘤之一[1],在妇科恶性肿瘤中其发病率仅次于宫颈癌和子宫内膜癌,死亡率最高,通常确诊时已为晚期并伴随转移[2-3]。据报道,卵巢癌患者的五年总体生存率约为30%[4]。卵巢癌的治疗方式主要以手术为主,辅以化疗的手段,而目前尚无卵巢癌的有效疗法,其发生和发展的分子机制尚未完全阐明。因此,从分子水平阐明影响卵巢癌发生、侵袭和转移等关键机制,将为确定卵巢癌治疗的新靶标以及对改善卵巢癌患者预后至关重要。
钙粘蛋白家族是一类细胞间粘附分子,主要分布在各种上皮组织中并介导异型细胞间的粘附[5]。N-cadherin是典型的钙粘着蛋白之一,在神经系统中广泛表达,而在其他正常细胞中则以低水平表达[6]。然而,近年来研究发现,N-cadherin在各种人类恶性肿瘤中异常表达,例如乳腺癌、前列腺癌、肺癌、肝癌等,其在癌细胞中的表达与肿瘤发生和转移密切相关[7-9]。另外,已有研究指出N-cadherin在多种卵巢癌细胞系如COLO316、ES2、SKOV3 和 WME-099 EBV中均异常高表达,通过靶向抑制N-cadherin使其功能丧失能够改善循环肿瘤细胞进而阻碍癌细胞的转移过程[10],推测N-cadherin对于判断卵巢癌的生物学行为具有重要的参考价值。然而,目前关于N-cadherin对卵巢癌发生发展的影响及其与病理机制的关系尚未完全明确。因此,本研究主要探讨沉默N-cadherin 对卵巢癌细胞生物学行为的影响,初步探究其作用机制,以期为卵巢癌的治疗提供新思路。
1 材料与方法
1.1 主要材料与试剂 卵巢癌细胞系SKOV3购自中国科学院典型培养物保藏委员会细胞库,病毒包装细胞株PT67购自上海索研生物科技有限公司。3%牛血清白蛋白、0.25%胰蛋白酶、100 U/mL青霉素、0.1 mg/mL链霉素及RPMI1640 培养液购自美国Sigma公司,Lipofectamine 3000试剂购自美国Invitrogen公司,Trizol试剂、Primescript RT reagent kit with gDNA Eraser试剂盒和SYBR Premix Ex Taq Ⅱ试剂盒购自日本Takara公司,CCK-8试剂盒、Transwell小室及免疫组织/细胞染色试剂购自上海碧云天生物研究所,RIPA 细胞裂解液、BCA蛋白定量试剂盒及增强型化学发光液ECL购自北京索莱宝科技有限公司,Matrigel 胶购自美国 BD 公司,兔抗N-cadherin、E-cadherin、MMP2、MMP9、MEK、ERK及鼠抗β-Actin购自美国 SantaCruz Biotech 公司,辣根过氧化物酶标记山羊抗兔IgG或山羊抗鼠 IgG抗体购自美国 Abcam公司。其他试剂均为国产分析纯或生物试剂。pEGFP-MSCVneo 质粒和 pMSCVneo/N-cadherin 质粒由上海生工生物科技有限公司构建。
1.2 实验方法
1.2.1 免疫组织化学染色检测N-cadherin在卵巢癌组织中的表达 收集2021年3月~2021年12月在我院进行手术切除的卵巢癌患者的卵巢癌组织和癌旁正常组织(距肿瘤组织≥1 cm,病例术前均有明确的病理诊断,且无化疗、放疗及免疫治疗史),所有组织均在手术切取后立即置于10%福尔马林溶液中固定。固定好后常规石蜡包埋,连续切片成5 μm,经脱蜡处理后,采用0.01 mol/L枸椽酸盐缓冲液修复抗原,3% 过氧化氢阻断内源性过氧化物酶,3%牛血清白蛋白(BSA)封闭 30 min,加入抗N-cadherin(1∶100)4℃孵育过夜,次日PBS 冲洗,加入HRP标记山羊抗兔IgG二抗(1∶1000)室温孵育30 min,PBS 冲洗,经显色、复染、脱水与透明后,中性树胶封片,光学显微镜采集图像并分析。
1.2.2 细胞培养 卵巢癌细胞系SKOV3复苏后,用含有10%胎牛血清(FBS)、100 U/mL青霉素与0.1 mg/mL链霉素的RPMI 1640 培养基置于37℃,5%CO2细胞培养箱中培养。每3 d换液1次,并传代1次。
1.2.3 RNAi 沉默 N-cadherin 基因的表达 将病毒包装细胞株PT67 接种至 6 孔培养板中,待其融合达 90%~95%时,参照Lipofectamine 3000 说明书用脂质体法将 pEGFP-MSCVneo 质粒和pMSCVneo/N-cadherin 质粒分别进行转染,收集病毒上清液。转染分为空白对照组、空载体组、siRNA-N-cadherin组。将对数生长期的SKOV3细胞以2×105个每孔接种至6孔板,37℃、5%CO2细胞培养箱中过夜孵育,将收集的 pEGFP-MSCVneo 病毒和 pMSCVneo/N-cadherin 病毒的上清分别感染空载体组和siRNA-N-cadherin组的细胞,然后用G418培养基进行维持培养。空白对照组不作任何处理,以RPMI 1640培养基继续培养。
1.2.4 CCK-8法检测各组细胞增殖能力 将转染后处于对数生长期的SKOV3细胞使用0.25%胰蛋白酶进行消化,并按照2×103个/孔接种于96孔板中,每组设置3个复孔,37℃、5%CO2细胞培养箱内继续培养,分别于12、24、48、72 h时每孔加入10 μL CCK-8试剂,在细胞培养箱内继续孵育2 h后,使用酶标仪在450 nm波长下检测各孔吸光度值。
1.2.5 细胞克隆形成实验检测各组细胞的克隆形成能力 将转染后处于对数生长期的SKOV3细胞用0.25%胰蛋白酶消化并吹打呈单细胞悬浮状态,以5×103个接种到培养皿中,轻轻晃动使细胞分散均匀,随后置于37℃、5%CO2细胞培养箱中培养14 d,出现细胞集落并用PBS 洗涤,加4%多聚甲醛固定菌落,用0.5%结晶紫染色15 min,流水冲洗,自然条件下干燥,在光镜下观察并计数细胞集落数目(将≥50个细胞聚集作为1个细胞集落)。
1.2.6 流式细胞术检测各组细胞周期分布 将转染后处于对数生长期的SKOV3细胞用PBS洗涤,预冷的75%乙醇重悬细胞,并在-20℃下固定24 h,PBS再次洗涤并重悬细胞,每组取 450 μL细胞悬液,加2 μL RNase后,37℃下孵育 10 min,接着加500 μL 碘化丙啶(PI),置于37℃下避光孵育30 min,过300目筛网,采用流式细胞仪检测细胞周期分布情况。
1.2.7 Transwell实验检测各组细胞的侵袭能力 在Transwell 小室的上室加50 μL 稀释的Matrigel胶,在37℃培养箱放置 40 min,将转染后的各组SKOV3细胞调整为 2×105个/mL,Transwell 小室的上室每孔加入 100 μL 细胞悬液,下室加入 600 μL含10%胎牛血清的RPMI 1640培养液培养至细胞贴壁,换无血清培养液培养12 h,放入37℃、5%CO2培养箱中孵育12 h,取出小室,弃培养液并轻轻擦去上室的细胞,4% 多聚甲醛固定 10 min,0.1%结晶紫染色10 min,PBS洗涤后封片,光学显微镜观察拍照,统计侵袭细胞数目。
1.2.8 细胞划痕实验检测各组细胞的迁移能力 在6孔培养板背后均匀画出横线,位置为每隔0.5 cm一道,每孔中加入转染24 h后的各组SKOV3细胞(每孔5×105个),第2天用移液枪枪头沿直尺垂直于细胞培养孔背后的横线划痕,PBS清洗细胞3次,洗去划痕下的细胞,然后加无血清RPMI 1640培养基,置于37℃、5%CO2细胞培养箱中继续培养,分别于0、24 h观察划痕愈合情况并拍照。
1.2.9 实时荧光定量PCR检测各组细胞N-cadherin、E-cadherin mRNA表达 使用Trizol法提取各组SKOV3细胞总RNA,NanoDrop2000分光光度计测定提取的RNA浓度与纯度。根据Primescript RT reagent kit with gDNA Eraser试剂盒说明进行操作将总RNA反转录为cDNA。采用SYBR Premix Ex Taq Ⅱ检测N-cadherin 、E-cadherin mRNA表达水平,以GAPDH 为内参。反应程序设置:95℃预变性3 min,95℃变性20 s,60℃ 退火 15 s,72℃ 延伸 45 s, 40 个循环。实验重复 3 次。qRT-PCR引物均由广州锐博生物公司合成,具体序列:N-cadherin 上游引物5′-GTGCCATTAGCCAAGGGAAT-3′,下游引物5′-GCGTTCCTGTTCCACTCATAG-3′;E-cadherin上游引物5′-GCTGGACCGAGAGAGTTTCC-3′,下游引物5′- CTGCATCTTGCCAGGTCCTT-3′; GAPDH上游引物5′-GTGAAGGTCGGAGTCAACG-3′,下游引物5′-TGGTGAAGACGCCAGTGGA-3′。采用2-ΔΔCt法分析N-cadherin、E-cadherin mRNA的相对表达量。
1.2.10 免疫细胞化学染色检测各组细胞中N-cadherin、E-cadherin、MMP2、MMP9蛋白表达 用0.25%的胰蛋白酶消化各组SKOV3细胞,将其制备为单细胞悬液,调整细胞浓度为4×105/mL,取0.5 mL接种在1 cm×1 cm 盖玻片上,贴壁后用PBS洗涤3次,在4%多聚甲醛固定20 min,3% 过氧化氢孵育后,加入3%BSA封闭 30 min,然后加入抗N-cadherin(1∶100)、E-cadherin(1∶100)、MMP2(1∶200)和MMP9(1∶200)4℃孵育过夜,次日PBS 冲洗,加入HRP标记山羊抗兔IgG二抗(1∶1000)室温孵育30 min,PBS 冲洗后DAB 显色,苏木素复染,脱水透明后中性树胶封片,光学显微镜采集图像并分析。
1.2.11 Western blot检测各组细胞中MEK、ERK蛋白表达 分别收集各组SKOV3细胞,加入预冷的RIPA 细胞裂解液充分裂解细胞,4℃条件下12000×g离心10 min取上清液,采用BCA法对蛋白质进行定量。分别取20 μg 每组蛋白样品,采用10%SDS-PAGE 电泳分离,将分离后的蛋白转移至PVDF膜,加5%脱脂奶粉室温封闭1 h后,分别加入兔抗MEK(1∶2000)、兔抗ERK(1∶3000)及内参鼠抗β-Actin(1∶3000),4℃下孵育过夜,次日TBST洗涤3次,每次10 min,加入辣根过氧化物酶标记山羊抗兔IgG或山羊抗鼠 IgG(1∶5000),室温下孵育2 h, TBST洗涤3次,滴加增强型化学发光液ECL,在Image Quant LAS 4000 mini下拍照并进行蛋白质条带灰度值的分析。

2 结果
2.1 N-cadherin 在卵巢癌组织中的表达 N-cadherin 阳性定位于细胞质,呈黄色或棕黄色颗粒,其在癌组织中的阳性率较癌旁正常组织中明显升高(71.85%vs12.46%,P<0.01),见图1。
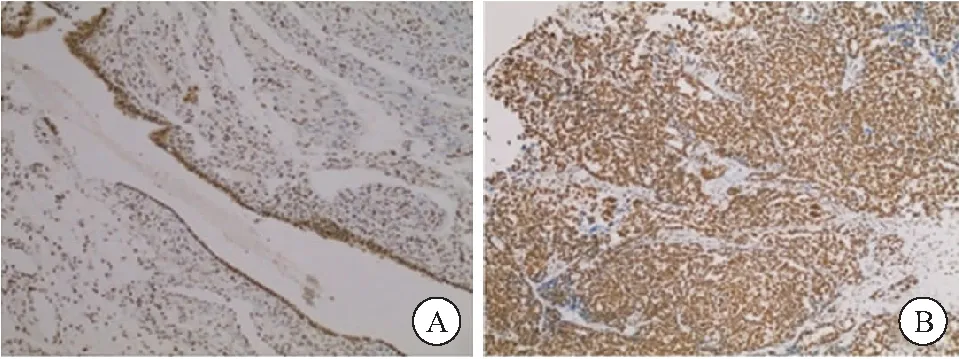
图1 卵巢癌组织和癌旁正常组织中N-cadherin表达免疫组织化学染色(100×)
2.2 沉默N-cadherin对卵巢癌细胞增殖的影响 CCK-8法检测结果显示,转染24、48和72 h后,siRNA-N-cadherin组的SKOV3细胞活力分别为(0.32±0.02)、(0.46±0.05)和(0.57±0.05),与空白对照组(0.53±0.05、0.73±0.13和0.85±0.24)和空载体组(0.51±0.07、0.74±0.11和0.85±0.30)比较显著下降(均P<0.05)(图2A)。提示沉默N-cadherin表达可降低SKOV3细胞增殖活力。细胞克隆形成实验结果显示,siRNA-N-cadherin组的SKOV3细胞克隆数目为(60.55±8.29)个,与空白对照组和空载体组[分别为(146.67±14.57)个、(139.27±16.45)个]比较显著下降(P<0.01)(图2B)。提示沉默N-cadherin表达可降低SKOV3细胞克隆形成能力。流式细胞术检测细胞分布情况,结果显示,空白对照组SKOV3细胞处于G1期、S期、G2期的细胞比例分别为(47.68±6.09)%、(35.29±5.76)%与(17.03±3.45)%,空载体组SKOV3细胞处于G1期、S期、G2期的细胞比例分别为(45.11±8.18)%、(38.29±4.01)与(16.60±2.23)%,siRNA-N-cadherin组G1期、S期、G2期的细胞比例分别为(34.01±3.89)%、(50.73±7.64)%与(15.26±2.03)%,siRNA-N-cadherin组较空白对照组和空载体组G1期细胞比例显著下降而S期显著增加(均P<0.01)(图2C)。提示沉默N-cadherin表达能够使SKOV3细胞发生S期阻滞。
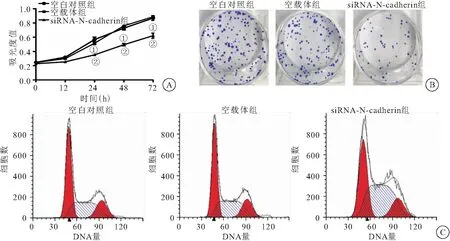
图2 沉默N-cadherin对卵巢癌细胞增殖的影响
2.3 沉默N-cadherin对卵巢癌细胞侵袭与迁移的影响 Transwell实验结果显示,siRNA-N-cadherin组SKOV3细胞侵袭数目为(201.56±23.60)个,较空白对照组和空载体组[分别为(522.78±52.58)个、(514.49±48.26)个]显著下降(均P<0.01)(图3A)。提示沉默N-cadherin表达可抑制SKOV3细胞的侵袭。细胞划痕实验结果显示,24 h后siRNA-N-cadherin组SKOV3细胞迁移数目为(320.77±28.79)个,较空白对照组和空载体组[分别为(605.81±57.71)个、(611.50±59.43)个]显著下降(均P<0.01)(图3B)。提示沉默N-cadherin表达可抑制SKOV3细胞的迁移。
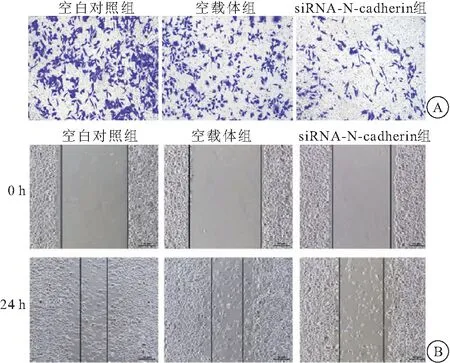
图3 沉默N-cadherin对卵巢癌细胞侵袭与迁移的影响
2.4 沉默N-cadherin对卵巢癌细胞上皮-间质转化的影响 qRT-PCR检测结果显示,siRNA-N-cadherin组N-cadherin mRNA与E-cadherin mRNA的相对表达量较空白对照组和空载体组N-cadherin mRNA相对表达量显著下降而E-cadherin mRNA相对表达量显著升高(均P<0.05),见表1。免疫细胞化学染色结果显示,空白对照组和空载体组细胞中可见到N-cadherin、MMP-2和MMP-9 蛋白阳性染色较强,阳性信号定位于细胞质,而siRNA-N-cadherin组中N-cadherin 、MMP2和MMP9蛋白的阳性染色较弱,与空白对照组和空载体组比较差异有统计学意义(均P<0.05);而siRNA-N-cadherin组中E-cadherin蛋白的阳性染色较空白对照组和空载体组显著增强(均P<0.05),见图4。提示沉默N-cadherin表达可抑制SKOV3细胞的上皮-间质转化。
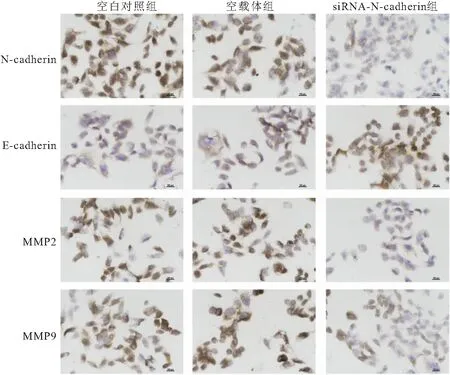
图4 免疫细胞化学染色检测各组SKOV3细胞中N-cadherin、E-cadherin、MMP2和MMP9蛋白表达(100×)
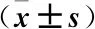
表1 各组N-cadherin、E-cadherin mRNA相对表达量
2.5 沉默N-cadherin对卵巢癌细胞MEK、ERK蛋白表达的影响 Western blot检测结果显示,siRNA-N-cadherin组MEK、ERK的蛋白表达量较空白对照组和空载体组显著下降(均P<0.05),见图5、表2。

图5 Western blot检测各组SKOV3细胞中MEK与ERK的蛋白表达
表2 各组MEK、ERK的蛋白表达量
3 讨论
卵巢癌是全世界最常见的妇科恶性肿瘤,由于早期缺乏明显的临床症状和特定的肿瘤标志物,大多数患者被诊断时已处于晚期[3,11]。目前,肿瘤标志物CA125的水平用于临床监测卵巢癌患者的预后,但其敏感性和特异性并不理想[12-13]。因此,探究卵巢癌的发生原因并确定精准的标志物以进行早期诊断,对于患者的预后评估和靶向治疗至关重要。
卵巢癌的增殖与转移过程涉及到多个过程包括癌基因的激活和抑癌基因的失活[14-15]。肿瘤的基因治疗较常规药物治疗具有特异性,目前,靶向基因治疗在卵巢癌的诊治中也备受瞩目。越来越多的证据表明,N-cadherin是上皮-间质转化的间充质标志物,其在许多肿瘤中呈阳性表达,其表达升高与肿瘤的侵袭与迁移密切相关[16-18]。Abufaraj等[19]评估了N-cadherin作为433例根治性膀胱切除术和双侧淋巴结清扫术的浸润性膀胱癌患者的预后生物标志物的作用,研究发现,浸润性膀胱癌患者中N-cadherin的阳性表达率约为40%。Kamikihara等[20]通过免疫组织化学抗生物素蛋白-生物素复合物法评估了146例术后胃癌患者肿瘤组织中N-cadherin的表达,发现N-cadherin的阳性表达与肿瘤的临床分期和恶性程度密切相关,实体瘤中N-cadherin表达明显上调。而关于N-cadherin在卵巢癌中的研究也已有报道,Po等[10]经检测卵巢癌细胞系中N-cadherin表达发现,其在多种癌细胞内呈现高表达,并影响循环肿瘤细胞捕获;于舒鸿等[21]研究指出在进展期卵巢癌、高级别浆液性癌及高级别卵巢癌中N-cadherin高表达,这与患者FIGO分期、组织学类型、肿瘤分级均显著相关。在本研究中,我们同样发现N-cadherin在卵巢癌组织中阳性表达明显升高。
为了进一步研究N-cadherin在卵巢癌中的作用,我们通过RNAi 沉默卵巢癌SKOV3细胞中 N-cadherin 表达,观察卵巢癌细胞生物学行为变化。qRT-PCR检测和免疫细胞化学染色结果均表明该干扰质粒能够有效抑制N-cadherin表达。在SKOV3细胞中沉默N-cadherin 表达后,细胞增殖活力明显下降,细胞集落形成数目减少,并使细胞发生S期阻滞,抑制细胞的迁移和侵袭能力。上皮-间质转化被认为是肿瘤细胞从非活动性上皮表型转变为迁移性间充质表型的过程,其共同特征是E-cadherin表达的丧失,同时伴有N-cadherin表达的上调[22]。MMP2和MMP9是基质金属蛋白酶家族的主要成员,可通过降解细胞外基质,血管基底膜和N-cadherin-catenin复合物来促进肿瘤细胞的侵袭、粘附以及转移。有研究表明,N-cadherin通过增加一些MMPs的表达来促进肿瘤转移[23-24]。本研究联合运用qRT-PCR和免疫组织化学法比较了 3 组细胞中 E-cadherin的表达情况,结果显示,沉默N-cadherin表达后E-cadherin的表达增加,MMP2和MMP9蛋白表达明显下降,说明上皮-间质转化过程受到抑制。
丝裂原激活蛋白激酶(MEK)/细胞外信号调节激酶(ERK)信号通路是多种细胞因子调控细胞生物学行为的主要途径,ERK被上游分子 MEK 的磷酸化激活后,可将信号从细胞膜传递至细胞核中,从而调节细胞的生命周期[25]。目前,研究表明MEK/ERK信号通路异常与多种肿瘤的发生、转移及恶化密切相关,该信号级联对肿瘤细胞间和肿瘤细胞内通讯至关重要,能够调节肿瘤细胞功能,如生长、存活和分化等,因此,该途径可作为干预和治疗包括卵巢癌在内的多种恶性肿瘤的潜在靶点[26]。本研究检测结果显示,在沉默N-cadherin表达的卵巢癌细胞中MEK与ERK的蛋白表达水平均明显下降,推测沉默N-cadherin可能抑制了MEK/ERK信号通路异常激活从而发挥了抑制卵巢癌生长与转移的作用。本研究进一步明确了N-cadherin在卵巢癌细胞恶性生物学行为的作用,为靶向N-cadherin治疗卵巢癌提供了新的科学资料。但本研究仅通过体外细胞实验研究了相关作用,而关于在体内沉默N-cadherin是否同样发挥积极作用及具体机制有待后续进行实验探究。
4 结论
本研究表明N-cadherin在卵巢癌组织中高表达,沉默卵巢癌细胞中N-cadherin表达可抑制细胞增殖、侵袭、迁移及上皮-间质转化,其机制可能与抑制MEK/ERK信号通路的异常激活有关。
猜你喜欢
生殖医学杂志(2022年10期)2022-10-19
智慧农业导刊(2022年7期)2022-04-21
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年1期)2022-02-28
中国药学药品知识仓库(2021年11期)2021-11-25
国际放射医学核医学杂志(2021年10期)2021-02-28
看世界·学术下半月(2020年7期)2020-09-10
中小学德育(2020年11期)2020-03-18
中国医药导报(2018年3期)2018-03-07
中国医学创新(2017年22期)2017-11-15
