
运动因子对机体代谢的调控作用及其机制研究进展
2022-08-15 07:06:14李秀儒傅力
中国运动医学杂志 2022年6期
李秀儒 傅力
天津医科大学医学技术学院康复医学系(天津 300070)
近些年来,运动调控机体代谢的生物学机制研究备受关注。现已证实,规律运动可预防和改善肥胖、2型糖尿病(type 2 diabetes mellitus,T2DM)、冠心病(coronary heart disease,CHD)、增龄性肌减症等多种代谢性疾病的发生和发展[1]。此外,运动还可提高大脑的认知功能,降低神经退行性疾病的发病风险[2]。运动过程中,机体器官和组织之间的交互作用(crosstalk)主要是由骨骼肌和脂肪等组织分泌的多种生物活性因子所介导。因此,这些因子被称为肌肉因子(myo⁃kines)和脂肪因子(adipocytes),特指运动过程中骨骼肌、脂肪组织所分泌的生物活性物质,骨骼肌和脂肪组织也因此被认为是具有内分泌功能的器官[3,4]。而其他器官和组织,如肝脏和心脏,也会在运动过程中释放代谢物和多肽等[5,6]。近些年来,随着运动医学研究的不断深入,运动因子(exerkines)已成为一个专有名词,特指机体器官和组织在运动过程中分泌的具有代谢调节作用的因子(代谢物及中间产物、多肽和RNA等物质)[7]。
1 运动因子的分泌
骨骼肌约占人体体重的40%~60%,是主要的运动器官和重要的内分泌器官。最初认为,骨骼肌分泌运动因子入血,通过血液循环发挥代谢调控作用[8]。随后研究发现,细胞外囊泡(extracellular vesicles,EVs)作为分子信号的重要载体,在介导运动效应中发挥重要作用。EVs 包括微囊泡(microvesicles,MVs)和外泌体(exosomes,EXs),是一种直径150~1000 nm 的膜性结构,几乎所有类型的细胞都可释放EVs[7]。运动过程中,骨骼肌、心脏、肝脏和脂肪组织等分泌的运动因子通过EVs 释放到细胞外,调控机体器官和组织之间的交互作用[9-11]。研究发现成年男性急性运动后血液循环中含多种microRNAs 的EVs 水平升高[7,12],长期有氧运动可提高人类和小鼠循环血中含多肽和核酸的EVs水平[11],这些含有运动因子的EVs可参与机体对运动的适应性调节、改善肥胖和T2DM 等代谢性疾病。值得注意的是,目前分离和纯化循环血液中含运动因子的EVs 仍较困难,常见的分离方法有超速离心法、超滤法和高分辨率质谱仪法等[13]。
2 运动因子的代谢调控作用及机制
2.1 白细胞介素6
白细胞介素6(interleukin 6,IL-6)是目前研究最为充分的运动因子之一。人体研究发现,急性运动可显著提高循环血IL-6 水平[14]。对健康[15,16]、肥胖个体[17,18],以及T2DM[17]患者进行急性运动干预后,骨骼肌和循环血中IL-6 水平立即增加。值得注意的是,运动诱导IL-6 基因表达和循环血水平在T2DM 和非T2DM肥胖个体之间无显著差别,提示T2DM 患者对运动具有良好的反应[17]。有趣的是,急性运动还提高人体脂肪组织IL-6转录水平,运动后脂肪组织IL-6基因表达迅速升高,并在恢复过程中仍维持较高水平[19]。
运动因子IL-6 作为运动过程中的能量传感器,可调节机体能量代谢[20,21]。一项随机对照试验发现,肥胖成年人经过12 周运动训练后,内脏脂肪组织(vis⁃ceral adipose tissue,VAT)的质量下降,而输注8 mg/kg IL-6受体抗体的受试者则未出现VAT质量下降,表明运动诱导的VAT 质量下降受IL-6 的调节[22]。Carey等研究表明,IL-6 可促进L6 肌管细胞葡萄糖转运体4(glucose transporter type-4,GLUT4)的膜转位,增加基础状态和胰岛素刺激下的葡萄糖摄取,并通过激活AMP 活化蛋白激酶(AMP-activated kinase,AMPK)增加脂肪酸氧化[23]。注射IL-6可促进小鼠跑台运动后胃肠胰高血糖素样肽-1(gastrointestinal glucagonlike peptide-1,GLP-1)的释放,从而增加胰岛素分泌,表明IL-6可调控骨骼肌-胰岛内分泌轴之间的交互作用[24]。此外,健康男性体内输注重组IL-6后进行3 h的功率自行车运动可降低肿瘤坏死因子-α(tumor ne⁃crosis factor-α,TNF-α)水平,提示IL-6 可缓解机体慢性炎症状态[25]。
2.2 成纤维细胞生长因子21
成纤维细胞生长因子21(fibroblast growth factor 21,FGF21)是一种主要来源于肝脏的新型运动因子,小鼠肝脏特异性敲除FGF21 基因后,循环血中FGF21水平显著下降[26]。10名健康青年男性进行低强度(55%VO2peak)和中等强度(75%VO2peak)的急性跑台运动后,循环血液中FGF21水平显著升高,且升高幅度与运动强度呈正相关[6]。Sabaratnam 等研究发现,14名肥胖个体和13 名T2DM 患者进行1 小时急性运动后,受试者骨骼肌FGF21 基因表达及循环血液浓度迅速增加,且在运动后3小时内保持上升趋势[17]。12周跑台运动可促进高脂饲料喂养的成年小鼠脂联素(adi⁃ponectin,ADPN)的分泌,同时诱导FGF21受体上调,从而抑制脂肪组织炎症[27]。但也有研究表明,3个月有氧运动和抗阻运动联合干预可降低肥胖女性循环血液中FGF21水平,其机制尚不清楚[28]。
多项研究表明运动因子FGF21可改善机体脂质代谢。临床研究发现,患T2DM 的肥胖个体使用FGF21类似物LY2405319 治疗后,血浆ADPN 和高密度脂蛋白胆固醇水平升高,空腹胰岛素、低密度脂蛋白胆固醇和甘油三酯水平降低、体重减轻[29]。动物实验也表明,人工合成的脂联素受体激动剂AdipoRon 可增加高脂饮食小鼠循环血液中FGF21 水平,并抑制白色脂肪组织(white adipose tissue,WAT)的脂解,降低血浆游离脂肪酸浓度,从而改善肥胖小鼠的全身胰岛素抵抗[30]。而FGF21 缺失则会导致脂质代谢异常,肝脏特异性敲除FGF21 的小鼠进行生酮饮食喂养后,出现脂肪肝、脂血症以及血酮降低的症状,表明FGF21在肝脏脂质氧化、甘油三酯清除和酮体生成中发挥重要作用[31]。
2.3 生长分化因子15
生长分化因子15 (growth differentiation factor 15,GDF15)是一种介导运动应激反应的细胞因子,与肌肉生长抑素(myostatin) 和卵泡抑素(fol⁃listatin,FST)同属于转化生长因子-β(transforming growth factor-β,TGF-β)超家族[32],通过GDNF家族受体α样(GDNF family receptor alpha like,GFRAL)受体发挥作用[33]。久坐不动的肥胖受试者进行12周有氧运动后,循环血GDF15水平升高,而患者腹部脂肪和总脂肪含量减少,提示GDF15 参与肥胖患者脂质代谢的调节[34]。小鼠进行60 分钟跑台运动后,肝脏GDF15的转录水平增加,表明GDF15 可能来源于肝脏[35]。Kleinert 等研究发现,健康年轻男性有氧耐力运动期间和运动后3小时内循环血GDF15水平均上升,但股动、静脉血GDF15 水平无显著差异,推测骨骼肌并不是GDF15 的主要来源[36]。然而,也有研究发现健康年轻男性进行1 小时急性运动后,骨骼肌GDF15 水平升高[37]。因此,运动过程中GDF15 的主要来源仍需进一步研究。
运动因子GDF15 通过调控机体能量代谢改善肥胖、T2DM、CHD 等疾病。在正常饮食和高脂饮食喂养的小鼠体内过表达GDF15,可改善小鼠糖耐量并减少摄食量[38]。Johnen等研究也发现,小鼠腹腔注射GDF15通过影响下丘脑摄食神经元减少摄食量,进而降低体重。上述表明GDF15可调节小鼠的摄食过程[39]。还有研究发现,对人的原代肌管细胞进行电脉冲刺激模拟急性剧烈运动或慢性中度运动,细胞培养液中GDF15水平升高,使用此培养基培养脂肪细胞可促进脂解作用[37]。此外,循环血GDF15 水平与中风、房颤、冠状动脉疾病和心肌梗死风险增加显著相关,提示GDF15 可作为心血管疾病潜在的生物标记物或治疗靶点[40]。
2.4 apelin
apelin 是一种新型运动因子,由13~36 个氨基酸组成[41],是孤儿G 蛋白偶联的APJ 受体的内源性配体,组成了apelin/APJ 系统,广泛存在于骨骼肌、心脏、肝脏、脂肪组织、胃肠道、脑、肾上腺等多种器官[42]。自发性高血压大鼠(spontaneously hypertensive rats,SHRs)进行长期游泳运动干预后收缩压降低,且ape⁃lin/APJ 的循环血液水平升高,提示运动训练对高血压的改善作用可能是通过提高apelin/APJ 水平实现的[43]。4周的低氧运动训练使肥胖小鼠体重下降,且小鼠骨骼肌apelin/APJ的基因和蛋白表达增加,表明肥胖小鼠体重的变化可能与骨骼肌中apelin/APJ 的水平有关[5]。Vinel 等发现,有氧运动显著提高年轻小鼠循环血液中apelin 的水平,而在老年小鼠中这种升高并不明显,推测可能与衰老过程中骨骼肌质量下降有关[44]。
根据靶点不同,运动因子apelin发挥的调控功能也不同。在骨骼肌中,apelin通过激活AMPK促进线粒体生物生成及线粒体自噬并降低骨骼肌炎症反应,存在于肌肉干细胞上的apelin 受体可促进干细胞增殖和分化并参与肌肉再生,而骨骼肌中apelin的表达随着年龄的增长而减少,提示apelin可能在肌减症发病过程中起调节作用[44]。在脂肪组织中,apelin通过促进脂肪细胞血管网络的生成,抑制了脂肪酸诱导的淋巴管和血管通透性增加,从而降低脂质堆积[45]。此外,apelin 是心血管疾病的一种生物标记物。研究表明,apelin 通过PI3K/Akt/FOXO3a 信号通路促进血管平滑肌细胞(vascular smooth muscle cells,VSMCs)迁移,从而在动脉粥样硬化的形成过程中发挥作用[46]。Zeng 等研究发现,apelin能抑制缺血心肌细胞凋亡和活性氧生成并降低乳酸脱氢酶漏出率及丙二醛含量,进而明显改善心肌缺血再灌注损伤引起的心功能减退[47]。
2.5 鸢尾素
鸢尾素(irisin)是一种由112 个氨基酸组成的糖基化蛋白,可与Ⅲ型纤维连接蛋白组件包含蛋白5(fi⁃bronectin type Ⅲ domain- containing protein 5,FNDC5)受体结合[48]。多种运动形式均可提高irisin水平,健康和患有代谢综合征的成年男性在进行高强度间歇运动(high-intensity interval exercise,HIIE)、持续中等强度运动(continuous moderate-intensity exer⁃cise,CMIE)和抗阻运动(resistance exercise,RE)后,循环血液中irisin水平立即升高,且对RE的反应最明显[49]。Bostrom 等研究发现,8名健康人进行10周的有氧耐力运动训练后,irisin的骨骼肌和循环血液水平均增加,提示irisin可能是一种来源于骨骼肌的运动因子[50]。然而,也有研究表明,健康和久坐少动男性进行12周有氧运动和抗阻运动联合训练后,循环血液中irisin水平并未改变,但骨骼肌FNDC5 表达显著增加[51]。因此,运动对irisin水平的具体调控机制还有待深入研究。
多项研究表明,运动因子irisin 可调节机体脂质代谢。胰岛素抵抗小鼠过表达FNDC5 可提高循环血液中irisin 水平,并使小鼠脂肪组织减少和体重下降,提示irisin 可改善胰岛素敏感性,增加能量消耗,并促进WAT 棕色化[50]。本课题组前期研究也发现,应激诱导蛋白Sestrin2通过FNDC5介导运动引起的皮下WAT棕色化过程,进而增加脂肪组织产热[52]。此外,细胞实验也表明irisin能增强3T3-L1细胞的脂解作用,是一种潜在促脂解因子[53]。irisin 不仅促进脂肪组织脂解产热,还在其他组织中发挥调节作用。Zhu 等研究发现,iri⁃sin可增加糖尿病小鼠内皮祖细胞(endothelial progen⁃itor cells,EPCs)的数量,改善血管内皮修复功能[54]。体外研究表明,irisin可与整合素aV/b5受体结合,促进骨细胞存活和硬化素的分泌,参与骨重建的局部调节[55]。
2.6 聚集素
聚集素(clusterin,CLU)于1983年被发现,是一种异源二聚体硫酸化糖蛋白,在肝、肾、睾丸等多种器官和组织中均有表达[56]。聚集素参与多种生理过程,如脂质运输、补体抑制及促进或抑制细胞凋亡等[57-59]。2021年,De 等发现,将运动28天小鼠的血液样本输入到安静对照组小鼠体内,对照组小鼠空间记忆力提高,神经系统炎症水平降低,且海马体中整体细胞的存活率增加。血浆蛋白质组学分析显示运动组小鼠血浆CLU 水平明显升高。为评估CLU 的功能,De 等给予急性脑炎和阿尔茨海默症小鼠模型静脉注射CLU,发现促神经炎性基因表达显著降低。此外,实验人员对20名具有轻度认知障碍的患者进行6个月运动干预后发现血浆CLU 水平升高[60],提示CLU 可能是一种具有抗炎作用的运动因子,有望成为运动防治神经退行性疾病的新靶点。
2.7 代谢中间产物和miRNAs
现有研究表明,一些运动过程中产生的代谢中间物和miRNAs 在机体代谢调控中发挥着重要作用。α-酮戊二酸(α-ketoglutaric acid,AKG)是三羧酸循环的一种关键代谢中间产物。研究发现,22 名志愿者进行60 分钟运动训练后,循环血液中AKG 水平显著升高,可能与运动引起的骨骼肌微损伤有关[61]。而自主跑轮运动和抗阻运动均可提高小鼠循环血液AKG 水平,且抗阻运动效果更明显。重要的是,AKG可促进肌肉肥大、棕色脂肪组织产热和白色脂肪组织脂解,其机制与2-氧戊二酸受体1(2-oxoglutarate receptor 1,OXGR1)刺激肾上腺释放肾上腺素有关[62]。此外,AKG 可以改善杜氏肌萎缩模型小鼠的肌萎缩症状,逆转糖皮质激素诱导的肌肉蛋白降解,提高小鼠肌肉力量和运动能力[63]。上述表明,AKG 可能是一种调节能量代谢和肌肉生长的重要运动因子。
miRNAs 同样可介导运动对机体代谢的调控作用。如4 周游泳运动对心肌梗塞小鼠产生保护作用,对小鼠血浆进行miRNA 分析,结果显示运动后miR-1192 显著升高。此外,心内注射agomiR-1192 也表现出类似的保护作用,而抑制miRNA-1192 后则消除了运动对心肌梗塞的保护效果。因此,miR-1192被认为是一种具有心肌保护功能的运动因子[64]。
3 总结与展望
运动因子作为骨骼肌、肝脏、脂肪等组织释放的机体代谢特异性调控因子,受到越来越多的关注。本文梳理了目前已知的运动因子IL-6、FGF21、GDF15、ape⁃lin、irisin的代谢调控作用及相关机制,总结了潜在运动因子CLU、AKG 和miR-1192 的最新研究结果。然而,运动因子和细胞外囊泡释放和摄取的分子机制,及其在组织交互作用中的潜在调节功能尚未完全阐明。因此,对运动因子的深入研究将为揭示运动调节机体代谢机制,科学选择运动频率、强度、时间和类型(fre⁃quency,intensity,time and type,FITT),合理制定个性化运动处方,将运动疗法应用于代谢性疾病的防治提供理论依据。
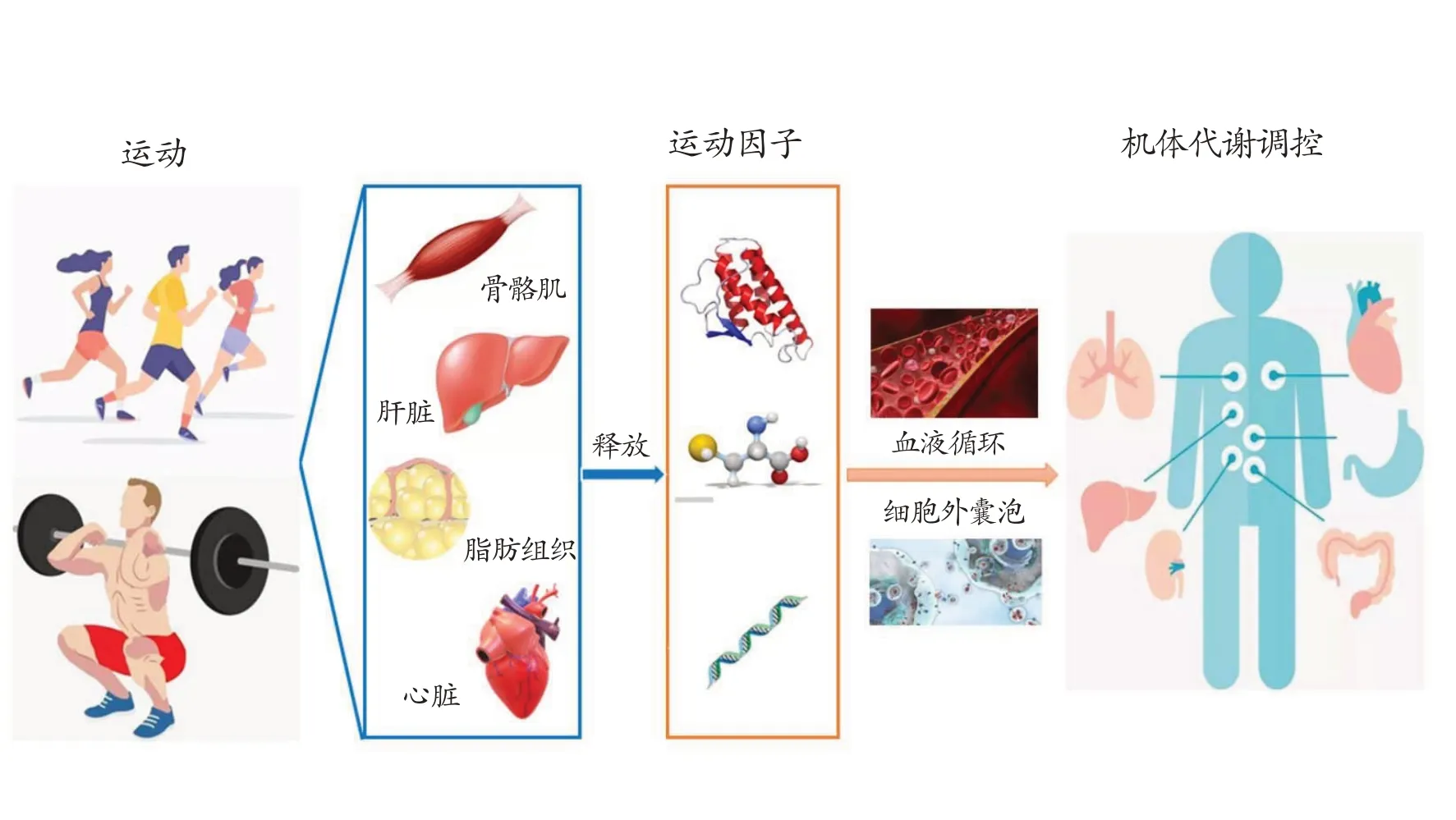
图1 运动因子作用示意图
猜你喜欢
中老年保健(2021年9期)2021-08-24 03:49:52
中老年保健(2021年7期)2021-08-22 07:40:46
昆明医科大学学报(2021年8期)2021-08-13 08:59:46
柴油机设计与制造(2018年3期)2018-10-13 01:45:08
中国铸造装备与技术(2017年3期)2017-06-21 11:33:37
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
医学研究杂志(2015年5期)2015-06-10 06:43:26
四川生理科学杂志(2014年2期)2014-02-28 14:09:09
河南医学研究(2014年2期)2014-02-27 14:51:37
