
贝氏小奥林螨连续饲养的草栖钝绥螨生命表参数和捕食能力
2022-08-13 08:08:20倪召红邹梦婷乙天慈郭建军金道超
昆虫学报 2022年7期
侯 飞, 倪召红, 邹梦婷, 朱 睿, 乙天慈, 郭建军, 金道超
(贵州大学昆虫研究所, 贵州省山地农业病虫害重点实验室, 农业农村部贵阳作物有害生物科学观察实验站, 贵阳 550025)
捕食性或寄生性天敌的成功饲养是生物防治计划的重要组成部分之一(Sanetal., 2020)。然而,大规模饲养天敌中品质退化是生物防治实践中常见的问题,而导致天敌品质和性能退化的重要原因之一是单一的营养来源(代平礼, 2005; 朱艳娟等, 2022)。单一的食物会使天敌缺乏某些营养物质而导致生活力和繁殖力衰退,且食物对天敌的影响表现相对滞后,可能在多代后才会显现(李会, 2011; 郭婉琳, 2016; Nemati and Riahi, 2020; 谢应强, 2020)。如捕食性天敌斯氏钝绥螨Amblyseiusswirskii以地中海粉螟Ephestiakuehniella卵饲养至第6代时,其种群参数显著降低,寄生性天敌烟蚜茧蜂Aphidiusgifuensis以烟蚜Myzuspersicae饲养至第12代时,其繁殖力、羽化率、僵蚜率均出现下降(Nguyenetal., 2014; 谢应强, 2020)。因此,食物对天敌品质和性能的影响需经过多代饲养后进行评估。
捕食性植绥螨已广泛应用于生物防治(Marcossietal., 2020)。草栖钝绥螨Amblyseiusherbicolus隶属于植绥螨科(Phytoseiidae),其异名为德氏钝绥螨A.deleoni(徐学农等, 2013)。该螨广泛分布于我国各地,国外分布于印度、伊朗、美国、新西兰等地(吴伟南等, 2009; 李洞洞, 2020),是一种多食性捕食螨,除捕食叶螨、跗线螨、瘿螨等害螨外(吴伟南等, 1991; Ferreiraetal., 2011; Rodríguez-Cruzetal., 2013; Duarteetal., 2015),还能捕食蓟马和粉虱等小型昆虫(Cavalcanteetal., 2015; Lametal., 2019, 2021),并在主要猎物缺乏时能以植物花粉为替代食物生存(Rodríguez-Cruzetal., 2013; Marcossietal., 2020),同时其对化学农药有一定抗性(Cordeiroetal., 2013; Reisetal., 2014),因而该螨是一种极具潜力的天敌。
目前,用于规模饲养捕食螨研究较多的为仓储粉螨,如甜果螨Carpoglyphuslactis、腐食酪螨Tyrophagusputrescentiae和椭圆食粉螨Aleuroglyphusovatus等(Nguyenetal., 2013; 郑苑等, 2019)。此外,江宏斌(2014)发现非仓储粉螨的温特螨科(Winterschmidtiidae)的贝氏小奥林螨Oulenziellabakeri可作为捕食性螨类的繁育猎物,该螨虽来自植物叶片但不以植物材料为食,不会对人体和植物健康带来威胁(江宏斌, 2014; Liu and Zhang, 2016)。基于此,贝氏小奥林螨是捕食螨的一种潜在的替代猎物,但贝氏小奥林螨作为单一食物连续饲养对捕食螨生活力和繁殖力的影响尚未见报道。
在生物防治计划中,对捕食者生活史和捕食作用进行准确的描述分析至关重要(Farhadietal., 2011)。生命表能全面描述昆虫种群的发育情况、存活率和繁殖力等,其中年龄-龄期两性生命表详细考虑了种群的龄期分化,并且包含雌、雄两性,能更加全面和准确地描述种群信息(Chi and Liu, 1985; 齐心等, 2019),而捕食功能反应是评价捕食性天敌控制害虫能力的重要方法(孙贝贝等, 2020)。因此,本研究采用年龄-龄期两性生命表和捕食功能反应测定草栖钝绥螨以贝氏小奥林螨为替代猎物连续多代饲养的生命表参数和捕食能力,以期为草栖钝绥螨天敌的扩繁利用提供理论基础。
1 材料与方法
1.1 供试虫源
草栖钝绥螨于2020年采自贵州大学南校区桂花树上(26°42′N,106°66′E,海拔1 083 m),用猎物贝氏小奥林螨饲养2个月(4~5个世代),使其达到相对稳定状态和实验所需虫量后开始实验;贝氏小奥林螨于2019年引自福州冠农生物科技有限公司,以酵母颗粒(安琪酵母股份有限公司)为食物建立种群;草栖钝绥螨和贝氏小奥林螨均在温度25±2℃,RH 85%±5%和光周期16L∶8D的人工气候箱内饲养。二斑叶螨Tetranychusurticae为贵州大学昆虫研究所在盆栽菜豆苗上保种12年的实验种群,饲养于温度25±5℃,RH 60%±10%,光周期12L∶12D的气候室中。
1.2 实验装置和仪器
1.2.1贝氏小奥林螨饲养装置:在正方形保鲜盒(18 cm×18 cm×8 cm)中放置厚度为2 cm的圆形海绵(d=9 cm),其上放置玻璃培养皿(d=9 cm),后加入酵母颗粒铺平培养皿,保鲜盒盖上切割9 cm×9 cm的方形孔,然后粘上200目的尼龙网用于空气流通。
1.2.2草栖钝绥螨群体饲养器:参照李颜等(2016)方法由两层有机玻璃组成饲养器,略作修改,底层为长×宽×厚=6 cm×5 cm×0.5 cm,中央切割直径4 cm圆孔并粘上200目黑色尼龙网,上层为长×宽×厚=6 cm×5 cm×0.3 cm作盖板。
1.2.3生命表实验小室:参照李颜等(2016)方法,略作修改,规格为底层为长×宽×厚=4 cm×3 cm×0.3 cm,中央切割直径2 cm圆孔并粘上200目黑色尼龙网,上层为长×宽×厚=4 cm×3 cm×0.2 cm作盖板。
1.2.4捕食功能反应观察小室:参照杨静逸等(2018)方法,略作修改,由3层4 cm×3 cm有机玻璃组成,底层厚度为0.2 cm,中层厚度为0.3 cm,中央有直径2 cm圆孔,底层与中层夹一张约3 cm长的干净菜豆叶,上层和中层规格相同,其圆孔上粘200目黑色尼龙网作为盖板,并用燕尾夹夹住两端。
1.2.5仪器:RXZ-260B智能型人工气候箱,宁波江南仪器厂;SMZ745体视显微镜,尼康仪器有限公司。
1.3 贝氏小奥林螨连续饲养的草栖钝绥螨两性生命表构建
参照Chi和Liu(1985)及Chi(1988)的方法构建年龄-龄期两性生命表。在草栖钝绥螨群体饲养器中放入适量缝纫线作为产卵基质,接入约60头草栖钝绥螨怀卵雌成螨,在25±2℃,RH 85%±5%和光周期16L∶8D下产卵24 h,将单粒卵挑入生命表实验小室,并添加足量猎物贝氏小奥林螨幼螨,并及时补充保证猎物充足并有剩余。未成熟期每天定时观察两次,成熟配对后每天观察一次,准确记录产卵量后将其移除(后代卵转移至新的生命表实验小室饲养至成螨,并收集至群体饲养器中饲养3~5 d,用于收集下一代卵),直至成螨死亡。从饲养器中挑出约60头怀卵雌成螨,所产的卵饲养至成螨,记为F1代。F1代成螨产卵用于组建F2代生命表,以上述方法依次收集F3代和F5代雌成螨所产的后代卵,分别组建F4代和F6代两性生命表。生命表实验均每隔5 d更换一次猎物。
1.4 贝氏小奥林螨连续饲养的草栖钝绥螨的捕食功能反应测定
参照吴坤君等(2004)的方法测定用贝氏小奥林螨饲养的草栖钝绥螨F2, F4和F6代雌成螨对二斑叶螨若螨和雌成螨的捕食功能反应,数据采用Holling圆盘方程Na=aTN/(1+aThN)拟合。F2, F4和F6代收集方法同1.3节,将收集的卵置于群体饲养器中饲养10 d后,将雌成螨挑至2.0 mL离心管饥饿24 h备用。二斑叶螨若螨和雌成螨密度设置为3, 6, 9, 12和15头/室,挑取相应二斑叶螨数量至观察小室后,接入1头经饥饿处理的草栖钝绥螨雌成螨,置于温度25±2℃,RH 85%±5%,光周期16L∶8D的人工气候箱内,24 h后观察记录其捕食情况,每处理重复5次。
1.5 数据分析
数据采用EXCEL 2019软件整理,导入两性生命表原始数据采用年龄-龄期两性生命表计算机程序TWOSEX-MSChart(Chi, 2021)软件进行计算分析得到主要参数,不同饲养代数数据差异显著性采用TWOSEX-MSChart程序的paired bootstrap test进行检验,参数的平均值与标准误用Bootstrap方法进行估计推断,其中Bootstrap次数为100 000次;作图采用Sigmaplot 14.0软件。捕食功能反应分析使用SPSS 23.0拟合方程,采用Duncan氏新复极差法进行差异显著性检验。
2 结果
2.1 贝氏小奥林螨连续饲养的草栖钝绥螨的生命表参数
2.1.1发育历期及寿命:贝氏小奥林螨连续饲养的草栖钝绥螨卵、第1若螨和第2若螨的发育历期以及产卵前期、总产卵前期和产卵期在F2, F4和F6代均无显著差异(P>0.05),但幼螨期、未成熟期(卵发育至成螨)、雌成螨和雄成螨寿命在各代之间存在显著差异(P<0.05)(表1)。F4和F6代幼螨期分别为1.01 d和1.02 d,显著长于F2代的0.98 d(P<0.05),而未成熟期有逐代延长的趋势。F2和F4代雌成螨寿命分别为22.30 d和21.35 d,而F6代显著下降为19.49 d(P<0.05),雄成螨寿命在F2代为21.60 d,而F4和F6代的分别显著下降至17.38 d和16.50 d(P<0.05),但不同代数雌成螨寿命均长于雄成螨。上述表明贝氏小奥林螨连续饲养会显著影响草栖钝绥螨成螨寿命。

表1 以贝氏小奥林螨为猎物的F2, F4和F6代草栖钝绥螨的发育历期和成螨寿命Table 1 Developmental duration and adult longevity of Amblyseius herbicolusin the F2, F4 and F6 generations reared on Oulenziella bakeri
2.1.2存活率和繁殖力:年龄-龄期存活率(Sxj)曲线(图1)描述以贝氏小奥林螨为猎物的F2, F4和F6代草栖钝绥螨存活率和发育速率的差异,反映出草栖钝绥螨不同发育阶段均有重叠现象。F2, F4和F6代草栖钝绥螨的未成熟期存活率分别为85.71%, 86.08%和88.00%,各代之间无显著差异(P>0.05)。在F2, F4和F6代,草栖钝绥螨卵存活率均为100.00%,其幼螨、第1若螨和第2若螨存活率在F2代分别为97.14%, 85.71%和81.43%,F4代分别为88.61%, 86.08和84.81%,F6代分别为92.00%, 88.00%和87.00%。同时,同代的雌、雄成螨的存活率最大值相同,F2, F4和F6代分别为42.86%, 43.04%和44.00%,雌成螨存活率最大值分别在第23.50, 20.50和18.50天时开始降低,雄成螨分别在第19.50, 18.50和15.50天时开始下降;在F2代时雄成螨与雌成螨的存活率下降速度一致,而F4代和F6代雄成螨存活率下降速度略快于雌成螨。上述表明贝氏小奥林螨连续饲养对草栖钝绥螨未成熟期存活率无显著影响。
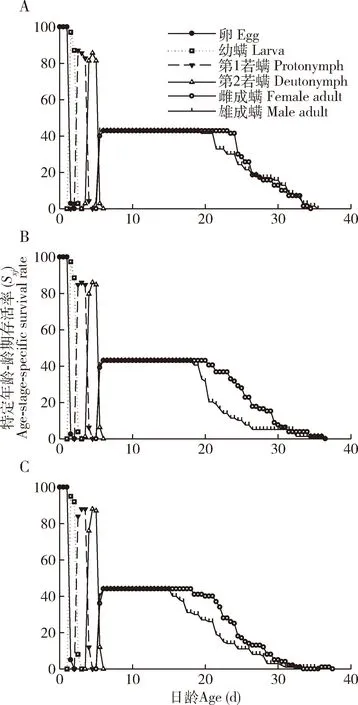
图1 以贝氏小奥林螨为猎物的F2(A), F4 (B)和F6(C)代草栖钝绥螨的特定年龄-龄期存活率(Sxj)Fig. 1 Age-stage-specific survival rate (Sxj) ofAmblyseius herbicolus in the F2 (A), F4 (B) andF6 (C) generations reared on Oulenziella bakeri
特定年龄存活率(lx)曲线(图2)反映以贝氏小奥林螨为猎物的F2, F4和F6代草栖钝绥螨从出生至死亡的变化过程。F2, F4和F6代种群存活率有两次较为明显的降低,且F2, F4和F6代草栖钝绥螨雌成螨繁殖力(fx5)和种群繁殖力(mx)曲线均呈先升高后下降的趋势,fx5和mx曲线均存在多次起伏波动。在F2和F6代,草栖钝绥螨雌成螨繁殖力fx5曲线的最大值分别在第10.0 和第11.0天时出现,分别为1.4667和1.2159粒/雌,但F4代的最大值在第7.5天时出现,为1.1765粒/雌。同时,其F2, F4和F6代的mx曲线最大值分别为0.73, 0.59和0.61。 F2, F4和F6代草栖钝绥螨的单雌产卵量无显著差异(P>0.05),分别为25.43±1.079, 24.37±1.38和24.45±0.79粒。

图2 以贝氏小奥林螨为猎物的F2(A), F4(B)和F6(C)代草栖钝绥螨的特定年龄存活率(lx)、特定年龄-龄期繁殖力(fx5)和种群特定年龄繁殖力(mx)Fig. 2 Age-specific survival rate (lx), age-stage-specificfecundity (fx5) and age-specific population fecundity (mx)of Amblyseius herbicolus in the F2 (A), F4 (B) andF6 (C) generations reared on Oulenziella bakeri
2.1.3种群参数:贝氏小奥林螨连续饲养的草栖钝绥螨在F2, F4和F6代的种群参数:总繁殖率(GRR)、净增殖率(R0)、内禀增长率(r)、周限增长率(λ)、平均世代周期(T)的平均值和标准误见表2。可以看出,在F2, F4和F6代,草栖钝绥螨各项种群参数均无显著差异(P>0.05)。

表2 以贝氏小奥林螨为猎物的F2, F4和F6代草栖钝绥螨的种群增长参数Table 2 Population growth parameters of Amblyseius herbicolus in the F2, F4 and F6 generations rearedon Oulenziella bakeri
2.2 贝氏小奥林螨连续饲养的草栖钝绥螨的捕食功能反应
F2, F4和F6代草栖钝绥螨对不同密度二斑叶螨若螨和雌成螨的捕食量见表3。结果表明,同一代数下,草栖钝绥螨对二斑叶螨若螨及雌成螨的捕食量均随着叶螨密度的增加而增大,但当叶螨密度达到一定程度后,其捕食量逐渐趋于饱和状态。在同一猎物密度下,F2, F4和F6不同代草栖钝绥螨对二斑叶螨若螨及雌成螨的捕食量均无显著差异(P>0.05)。同时,同一代草栖钝绥螨对二斑叶螨若螨的捕食量高于对二斑叶螨雌成螨的捕食量,因此,草栖钝绥螨对二斑叶螨若螨的捕食作用强于雌成螨。

表3 以贝氏小奥林螨为猎物的F2, F4和F6代草栖钝绥螨对二斑叶螨若螨和雌成螨的平均日捕食量Table 3 Average daily consumption of Amblyseius herbicolus in the F2, F4 and F6 generations reared onOulenziella bakeri to nymphs and female adults of Tetranychus urticae


表4 以贝氏小奥林螨饲养的F2, F4和F6代草栖钝绥螨对二斑叶螨若螨和雌成螨的捕食功能反应方程及参数Table 4 Predation functional response and parameters of Amblyseius herbicolus in the F2, F4 and F6 generations rearedon Oulenziella bakeri to nymphs and female adults of Tetranychus urticae
表中数据为平均值±标准误;同行数据后不同字母表示经paired bootstrap检验差异显著(P<0.05)。Data in the table are means±SE, and means followed by different letters within a row are significantly different according to paired bootstrap test (P<0.05).
3 结论与讨论
连续大规模饲养天敌时,由于稳定的饲养环境、营养来源单一和近亲繁殖等因素会影响天敌品质和性能,常表现为种群发育整齐度、存活率、繁殖力、寿命、捕食或寄生能力等出现不同程度的衰退现象(代平礼, 2005; Guzmán-Larraldeetal., 2014; 岳方正, 2016; 谢应强, 2020; Ghaemmaghamietal., 2021)。目前,寄生性天敌连续多代饲养后的种群退化现象报道较多(岳方正, 2016; 谢应强, 2020; 朱艳娟等, 2022),但在捕食螨中研究相对较少,仅见商业化加州新小绥螨Neoseiuluscalifornicus和斯氏钝绥螨有研究报道(Nguyenetal., 2014; Khanamanietal., 2017; Nemati and Riahi, 2020)。
天敌昆虫在单一食物饲养情况下,发育历期通常随着饲养代数的增加逐渐延长(岳方正, 2016; Nemati and Riahi, 2020; 朱艳娟等, 2022)。本研究发现,F2, F4和F6草栖钝绥螨的发育历期逐渐延长,再次印证了该结论。本研究还发现,草栖钝绥螨雌成螨在6代、雄成螨在4和6代的寿命显著缩短(表1)。该结果也再次印证了单一食物饲喂天敌,其寿命会随饲养代数增加而显著缩短(谢应强, 2020)。
食物营养能否满足天敌生存和繁殖十分重要。本研究结果表明,贝氏小奥林螨作为单一替代食物,可以满足草栖钝绥螨的生存和繁殖需求。本研究中,F2, F4和F6代草栖钝绥螨的未成熟期存活率及单雌产卵量均无显著差异(图1和2)。此结果与Nguyen等(2014)利用丰年虾Artemiafranciscana卵连续饲养斯氏钝绥螨,其后代未成熟期存活率差异不显著,Khanamani等(2017)利用杏花粉连续饲养加州新小绥螨,对其后代的单雌产卵量无显著影响的结果类似。表明贝氏小奥林螨能够满足草栖钝绥螨的生存和繁殖需求。
内禀增长率是衡量某种群在特定的环境条件下的消长趋势和适合度的一个重要指标,周限增长率是在特定环境中某种群内平均每一个个体能产生的后代数(Farhadietal., 2011)。当种群内禀增长率(r)大于0,周限增长率(λ)大于1时,种群数量呈增长趋势(刘向东, 2016; 赵远鹏等, 2018)。本研究发现,贝氏小奥林螨饲养的F2, F4和F6代草栖钝绥螨r和λ值分别在0.19~0.20和1.21~1.22/d之间,同时发现各代之间的种群参数无显著差异(表2),表明贝氏小奥林螨饲养的草栖钝绥螨种群数量呈增长趋势。表明单一食物连续多代饲养天敌不会有显著影响其主要种群参数。该结果与Song等(2019)利用添加地中海粉螟卵人工饲料饲养斯氏钝绥螨,Nguyen等(2014)利用丰年虾卵饲养斯氏钝绥螨,Khanamani等(2017)利用杏花粉饲喂加州新小绥螨所得结果类似。
目前,捕食功能反应仍然是评价捕食性天敌对害虫控制能力大小的基本方法之一(段雪莹等, 2021)。有研究表明,草栖钝绥螨对柑橘全爪螨Panonychuscitri、紫红短须螨Brevipalpusphoenicis和金雀花植物上蓟马Sericothripsstaphylinus的功能反应类型符合Holling Ⅱ型(高平等, 1990; Reisetal., 2007; Lametal., 2021)。本研究得出F2, F4和F6代草栖钝绥螨对二斑叶螨若螨和雌成螨的捕食功能反应均符合Holling Ⅱ型,可见连续饲养并未影响草栖钝绥螨的功能反应类型。此外,本研究在同一猎物密度下,发现F2, F4和F6代草栖钝绥螨对二斑叶螨若螨及雌成螨的捕食量均无显著差异(表3),这与Nguyen等(2014)用人工饲料饲养斯氏钝绥螨之F1代和F6代对西花蓟马Frankliniellaoccidentalis1龄若虫的捕食量无显著差异的结果一致。捕食功能反应参数中,瞬时攻击率(a)和处理时间(Th)是评价捕食者对猎物捕食作用大小的方法之一,其比值大小能直接反映捕食者对猎物的控害能力(贾静静等, 2019; 焦进卫等, 2021)。本研究结果发现,F4和F6代草栖钝绥螨对二斑叶螨若螨、F6代对二斑叶螨雌成螨的捕食能力(a/Th)明显下降(表4),该结果与Castagnoli和Simoni(1999)研究加州新小绥螨以粉尘螨Dermatophagoidesfarinae在实验室内连续饲养4年后与野外品系相比,其捕食能力有下降趋势,以及与Nguyen等(2014)利用丰年虾卵和人工饲料饲养斯氏钝绥螨至第6代,其适合度出现不同程度下降的结果类似,即随着饲养代数的增加捕食螨的捕食能力出现下降趋势,究其原因,可能是捕食螨的食物适应性下降所致。
总之,以贝氏小奥林螨为替代猎物的草栖钝绥螨能正常发育和繁殖,本研究饲养代数内,其生命表参数和捕食量未出现明显下降现象,其种群数量呈增长趋势。因此,贝氏小奥林螨可作为草栖钝绥螨扩繁饲养的替代猎物,但需进一步深入研究贝氏小奥林螨长期饲养草栖钝绥螨后可能出现捕食能力下降的问题及相应的解决措施,以促进草栖钝绥螨在生物防治中应用。
猜你喜欢
人类学学报(2023年5期)2023-04-29 19:50:21
青少年科技博览(中学版)(2022年9期)2022-11-01 08:22:44
海洋渔业(2021年1期)2021-03-18 01:53:34
水生生物学报(2021年1期)2021-02-04 06:39:32
第二课堂(小学版)(2019年7期)2019-07-16 05:26:15
猪业科学(2018年4期)2018-05-19 02:04:52
运动(2017年1期)2017-09-03 10:39:20
金色少年(奇趣科普)(2017年1期)2017-03-03 07:05:38
中国科技信息(2016年12期)2016-08-29 01:08:47
科技视界(2015年26期)2015-09-11 15:21:45

- 昆虫学报的其它文章
- Morphological differentiation in the Asian honey bees (Apis cerana) in China
- 我国余甘子新害虫锡兰玻壳蚧的形态及生物学特性
- 施药后不同时间玉米田中常用农药残留对玉米螟赤眼蜂的影响
- Inhibition of glucose oxidase gene decreases the resistance of Mythimna separata (Lepidoptera: Noctuidae) larvae to Bacillus thuringiensis infection
- 枣食芽象甲化学感受蛋白PyasCSP4的基因克隆、表达及配体结合特征
- RNAi介导的GdHsp60和GdHsp70基因沉默对沙葱萤叶甲幼虫抗寒性的影响