
生草对果园生态系统服务功能的影响: 全球数据整合分析研究*
2022-08-13 05:58高晓东赵西宁于流洋何娜娜
中国生态农业学报(中英文) 2022年8期
呼 盼, 高晓东, 赵西宁, 于流洋, 何娜娜
(1. 西北农林科技大学旱区农业水土工程教育部重点实验室 杨凌 712100; 2. 西北农林科技大学水利与建筑工程学院 杨凌712100; 3. 中国科学院水利部水土保持研究所 杨凌 712100)
果园作为一种农业生态系统, 其生态系统服务功能受到人类管理措施的巨大影响。通常, 果农为了获得更高的产量, 提升果园的供给服务能力, 会采取一系列田间管理措施, 比如去除行间杂草进行清耕、投入大量的化肥农药等。但是这种传统集约经营模式的长久实施会造成水土流失、固碳能力减弱、硝酸盐污染等环境问题, 严重损害了果园调节和支持方面的生态系统服务功能。果园生草可以增加生物多样性和有机物投入, 减少无效蒸发损失和水土流失, 是一种调控果园生态系统服务功能的重要途径, 有利于构建良好的果园生产体系、改善果园环境、提高果园生产力。因此, 探明果园生草对生态系统服务功能的影响对探寻果园可持续管理策略具有重要科学价值。
欧美日等发达国家较早意识到了果园生草的重要性, 果园生草已经成为其果园建设的主流模式。在西方发达国家, 生草果园面积已占果园总面积的60%~75%以上, 在有的国家推广面积甚至超过了90%。而我国果园生草面积不足10%。我国相关研究起步较晚, 20世纪80年代才开始借鉴国外先进经验开展果园生草技术研究。到1998年, 农业部中国绿色食品发展中心提出将果园生草作为绿色果品生产技术体系在全国推广。但在农耕历史、农业现状等经济、社会、自然诸多主客观因素长期影响下, 生草果园面积仍然较少, 近20年我国有针对性地开展了大量果园生草特色鲜明的研究, 取得了丰硕的研究成果。例如刘业萍等研究发现生草可显著增加土壤中总有机碳和可溶性有机碳含量, 进而显著增加了土壤中微生物数量, 并不同程度地改变了细菌和真菌的丰富度和多样性; 郭晓睿等综述了果园生草对中国果园土壤肥力和生产力的影响, 发现果园生草可以提高土壤有机质、碱解氮、速效磷含量, 降低土壤容重; 陈久红等研究发现果园生草改善了果园生态环境, 促进了叶片光合作用, 改善了果实品质, 为果树高产优质创造了条件。众多研究表明, 果园生草通过增加生物量和土壤碳固存在调控气候方面发挥着至关重要的作用, 并且通过缓冲极端气候和阻断直接辐射来适应气候变化, 还可以通过提供多种栖息地和改善小气候, 保护生物多样性, 提升功能多样性、养分循环和土壤理化性质。尽管大量案例研究评估了果园生草的生态系统服务, 但是由于果园类型、生草种类以及外界环境存在明显差异, 导致研究结果呈现显著的异质性。虽然已有相关文献综述了果园生草生态系统服务功能方面的进展, 但无法系统、整体、定量地评价果园生草的生态系统服务功能。整合分析(meta-analysis, 又称为元分析)作为一种基于大量数据的定量分析方法, 为系统和整体地了解果园生草的生态系统服务功能提供了重要手段, 然而目前关于全球果园生草生态系统服务功能评价的整合分析研究较少。
本研究采用整合分析方法, 定量评估了全球果园生草的生态系统服务功能, 主要包括供给(产量和品质)、调节(土壤水分、温度、有机碳)以及支持(全氮)等服务功能, 并分析了不同果园类型、生草种类以及环境条件下生态系统服务功能的异质性, 以期通过系统和整体地认识生草对生态系统服务功能的影响, 为果园可持续管理提供科学依据。
1 材料与方法
1.1 数据来源
本研究以“果园生草” “果园间作” “土壤管理”等关键词在Web of Science和中国知网数据库中对1996−2020年发表的果园生草方面的文献进行检索, 初步筛选出果园生草对生态系统服务功能影响的相关文献。
通过详细分析文献的研究目的、研究方法和研究结果, 结合本次研究目的, 解决数据点的非独立性问题, 设立了3条数据的搜集筛选标准: 1)试验必须同时包含清耕组(对照组)和果园生草处理组(试验组); 2)在野外试验中, 试验组和对照组的试验场所、土壤质地以及气候特征等需保持一致; 3)试验组和对照组中与果园生草相关的变量的平均值以及样本量能够直接从图、表或者文字中进行提取, 或者以上数据可以直接从文献中通过计算获得。
1.2 数据整理和分类
经过进一步文献筛选, 最终确定了符合相关标准的文献共计118篇。结合筛选出的文献, 利用Excel 2010 对筛选到的文献进行建库, 如果文献以图片显示数据, 则使用GetData Graph Digitizer 2.24 对图片数据进行数字化。数据库主要是生态系统服务功能指标数据, 包括果实产量和品质、土壤有机碳、全氮、土壤温度和土壤水分。遵循联合国千年生态系统服务分类方案分类, 将生态系统服务功能细化为: 供给服务功能、调节服务功能、支持服务功能及文化服务功能(https://www.millenniumassessment.org/zh/Framework.html)。依据上述功能定义, 把果实产量与品质作为供给服务, 土壤水分、温度及有机碳作为调控服务, 土壤全氮作为支持服务。
除此之外还提取了作者信息、试验地点及其经纬度、土壤利用类型、土壤质地、果园生草种类与年限等内容。根据文献中提取到的相关试验信息进行归纳分组, 整理得到以下影响因素: 海拔、气候类型、生草种类、果园类型。海拔和气候能够显著影响降雨、气温、辐射等环境因素以及果树和草种分布, 生草种类和果园类型本身是影响生态系统服务功能的主要因素, 因此, 选择上述4个因素进行分析。参考Kuyah等研究, 海拔分为低海拔(1200 m以下)和高海拔(1200 m以上); 根据文献中提取的气候类型, 分为干旱区、湿润区和半湿润区; 生草种类分为豆科(Leguminosae)类、禾本科(Gramineae)类和杂草。果园类型分为蔷薇科(Rosaceae)、芸香科(Rutaceae)、鼠李科(Rhamnaceae)、葡萄科(Vitaceae)、橄榄科(Burseraceae)和石榴科(Punicaceae)等。
1.3 整合分析方法
响应比(response ratios, RR)是用于评估试验处理对某一变量影响程度的统计学指标, 对于某一指定变量, RR为试验组()与对照组()的平均值之比, 计算公式如下:

本文采用效应比值RR的自然对数计算果园生草对生态系统服务功能的效应:
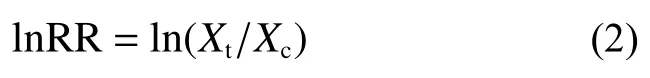
LnRR=0, 生草并未引起试验组和对照组变量之间的差异; LnRR<0, 说明果园生草对所关注变量产生了负效应; LnRR>0, 则说明果园生草对所选变量产生了正效应。
通过使用MATLAB编程语言应用10 000次迭代, 使用Bootstrapping方法估计不同类别变量的LnRR加权均值的95%置信区间。整合分析中, 计算指标总体的平均值与单个个体的观测值权重密切相关, 在计算单个观测值的权重时大多数研究的标准差缺失, 但样本量是可用的。因此, 根据描述的样本大小, 使用以下公式计算LnRR的权重:

式中:为单个观测结果的权重,和分别为试验组和对照组的样本容量。
2 结果与分析
如图1所示, 与对照区相比, 果园生草显著提高了果实产量, 增加土壤含水量、土壤有机碳及全氮含量; 生草果园的果实产量、可溶性固形物含量、土壤有机碳含量、土壤含水量和全氮含量分别增加20.7%、5.1%、24.7%、8.1%和15.6%, 表明生草对果园这些生态系统服务功能均具有显著的正效应。但对果实品质中的可滴定酸和土壤温度产生了明显的负效应, 分别较对照组降低10.8%和10.6%。
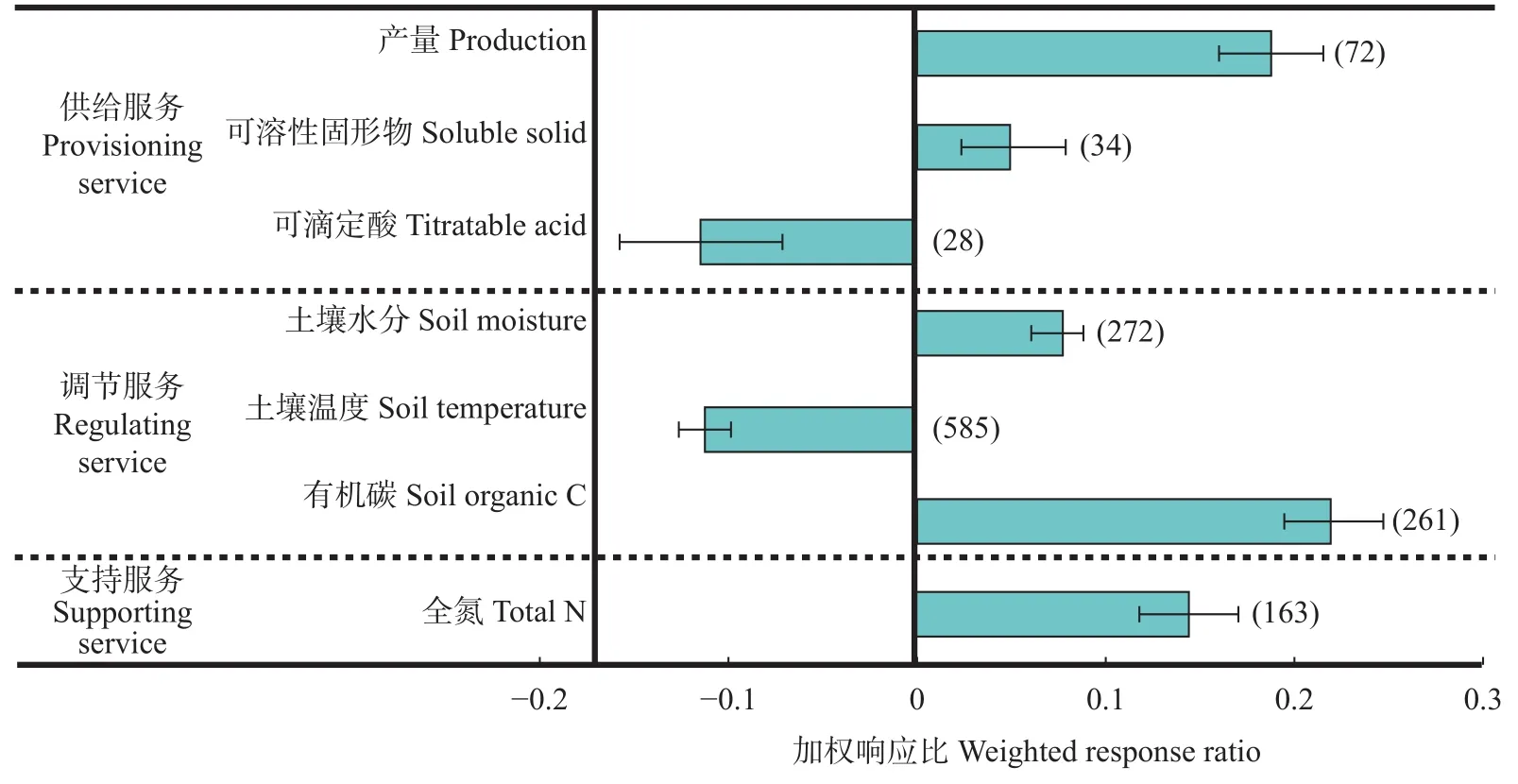
图1 生草对生态系统服务功能的总体影响Fig.1 General effects of sod-culture on orchard ecosystem services function
2.1 生草对果园供给服务功能的影响
2.1.1 果实产量
不同果园类型、生草种类、气候和海拔条件下生草对果实产量的影响如图2所示。与果园清耕相比, 各立地条件下生草对果实产量的影响均为正效应。就果园类型而言, 橄榄科、石榴科、芸香科和蔷薇科的平均效应值分别为0.092 (置信区间为0.021~0.164)、0.247 (置信区间为0.199~0.296)、0.247(置信区间为0.193~0.300)和0.173 (置信区间为0.132~0.215)。就生草种类而言, 豆科效应值最高, 平均为0.206 (置信区间为0.151~0.261), 高于杂草的0.154 (置信区间为0.100~0.206)和禾本科的0.171 (置信区间为0.124~0.218)。就气候而言, 湿润区平均效应值为0.225 (置信区间为0.130~0.319), 高于半湿润区的0.173 (置信区间为0.141~0.205)和干旱区的0.201 (置信区间为0.147~0.254)。就海拔而言, 高海拔平均效应值为0.243 (置信区间为0.028~0.461), 高于低海拔的0.176 (置信区间为0.138~0.213)。
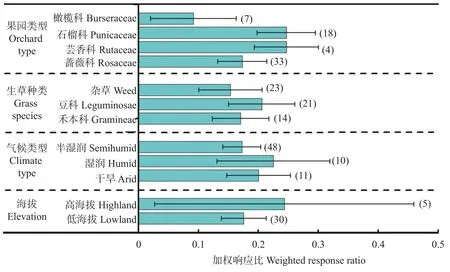
图2 不同条件下生草对果园产量的影响Fig.2 Effects of sod-culture on yield of orchard under different factors
2.1.2 果实品质
选取可溶性固形物和可滴定酸两个指标反映果实品质。不同果园类型、生草种类、气候和海拔下果实可溶性固形物分析结果如图3所示。就果园类型而言, 生草对葡萄科、芸香科和蔷薇科果园可溶性固形物均为正效应, 平均分别为0.012 (置信区间为0.004~0.020)、0.132 (置信区间为0.052~0.212)和0.043 (置信区间为0.013~0.073), 但鼠李科果园为负效应, 效应值为−0.066 (置信区间为−0.104~−0.028)。不同生草种类的效应值相当, 分别为: 杂草0.048 (置信区间为−0.040~0.139)、豆科0.070 (置信区间为0.034~0.106)、禾本科0.067 (置信区间为0.047~0.088)。就气候类型而言, 湿润区效应值平均为0.117 (置信区间为0.014~0.220), 高于半湿润区的0.053 (置信区间为0.037~0.070), 但干旱区为负效应, 平均为−0.058(置信区间为−0.091~−0.025)。就海拔而言, 高海拔为负效应, 平均为−0.045 (置信区间为−0.076~−0.015), 而低海拔为正效应, 平均为0.053 (置信区间为0.033~0.074)。
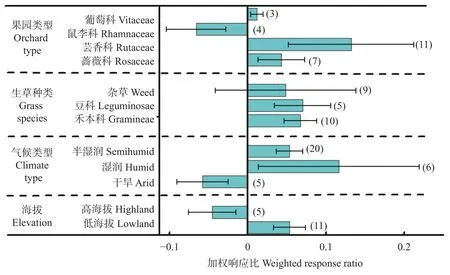
图3 不同条件下生草对果实可溶性固形物影响Fig.3 Effects of sod-culture on soluble solid in fruit under different factors
不同果园类型、生草种类、气候和海拔下果实可滴定酸分析结果如图4所示。与果园清耕相比, 生草对果实可滴定酸整体上为负效应。就果园类型而言, 芸香科的效应值平均为−0.290 (置信区间为−0.347~−0.232), 低于蔷薇科的−0.087(置信区间为−0.160~−0.012)。就生草种类而言, 杂草的效应值平均为−0.049 (置信区间为−0.146~0.049), 高于豆科的−0.166 (置信区间为−0.222~−0.111)和禾本科的−0.172 (置信区间为−0.233~−0.111)。就气候类型而言, 半湿润区的效应值为−0.147 (置信区间为−0.180~−0.119), 但干旱区表现出正效应, 达0.050 (置信区间为0.001~0.098)。就海拔而言, 高海拔为正效应, 达0.057 (置信区间为0.019~0.096), 而低海拔为明显负效应, 平均为−0.158 (置信区间为−0.183~−0.134)。

图4 不同条件下生草对果实可滴定酸的影响Fig.4 Effects of sod-culture on titraTableacid in fruit under different factors
2.2 生草对果园调节服务功能的影响
2.2.1 土壤水分
不同立地条件下生草对果园土壤水分影响的效应值如图5所示。与清耕相比, 生草对果园土壤水分影响为显著正效应。就果园类型而言, 鼠李科果园的正效应平均值达0.180 (置信区间为0.157~0.202), 高于蔷薇科果园的0.071 (置信区间为0.060~0.083)。就生草种类而言, 杂草和禾本科的效应值分别为0.139 (置信区间为0.006~0.216)和0.131 (置信区间为0.117~0.146), 高于豆科的0.057 (置信区间为0.037~0.077)。就气候类型而言, 半湿润区效应值为0.068 (置信区间为0.056~0.080), 低于干旱区的0.171(置信区间为0.152~0.189)。就海拔而言, 高海拔表现为负效应, 平均效应值为−0.001 (置信区间为−0.021~0.019), 而低海拔为显著正效应, 达0.104 (置信区间为0.082~0.126)。

图5 不同条件下生草对果园土壤水分的影响Fig.5 Effects of sod-culture on soil moisture in orchard under different factors
2.2.2 土壤温度
不同立地条件下生草对果园土壤温度影响的效应值如图6所示。与对照组相比, 生草对果园土壤温度的影响均表现出负效应。就果园类型而言, 鼠李科和蔷薇科效应值分别为−0.107 (置信区间为−0.142~−0.073)和−0.113 (置信区间为−0.129~−0.097), 二者相近。就生草种类而言, 杂草、豆科和禾本科的效应值分别为−0.123 (置信区间为−0.161~−0.084)、−0.120 (置信区间为−0.142~−0.098)和−0.096 (置信区间为−0.116~−0.078), 三者之间的差异也较弱。就气候类型而言, 半湿润区效应值平均为−0.098 (置信区间为−0.109~−0.087), 明显高于干旱区的−0.158 (置信区间为−0.211~−0.104)。就海拔而言, 高海拔的效应值平均为−0.197 (置信区间为−0.277~−0.115), 低于低海拔的−0.077 (置信区间为−0.084~−0.071)。

图6 不同条件下生草对果园土壤温度的影响Fig.6 Effects of sod-culture on soil temperature in orchard under different factors
2.2.3 土壤有机碳
不同立地条件下生草对果园土壤有机碳影响的效应值如图7所示。与果园清耕相比, 生草对果园土壤有机碳影响整体上为正效应。就果园类型而言, 芸香科效应值平均达0.461 (置信区间为0.239~0.685), 高于鼠李科的0.316 (置信区间为0.231~0.393)和蔷薇科的0.276 (置信区间为0.248~0.303)。就生草种类而言, 杂草效应值平均为0.201 (置信区间为0.139~0.265), 低于豆科的0.294 (置信区间为0.257~0.331)和禾本科的0.299 (置信区间为0.248~0.350)。就气候类型而言, 干旱区效应值平均为0.310 (置信区间为0.207~0.410), 高于半湿润区的0.221 (置信区间为0.199~0.244)和湿润区的0.150 (置信区间为0.078~0.222)。就海拔而言, 低海拔效应值为0.226 (置信区间为0.196~0.253), 高于高海拔的0.096 (置信区间为−0.201~0.390).
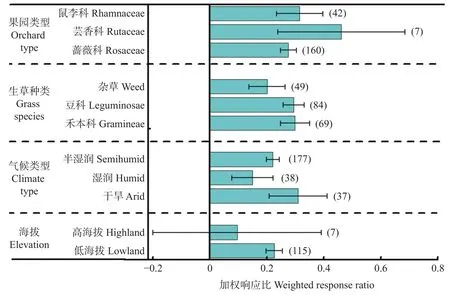
图7 不同条件下生草对果园土壤有机碳含量的影响Fig.7 Effects of sod-culture on soil organic carbon content in orchard under different factors
2.3 生草对果园支持服务功能的影响
选取全氮作为土壤支持服务功能的指标, 不同立地条件下生草对果园土壤全氮含量影响的效应值如图8所示。与果园清耕相比, 生草整体上提高了果园土壤全氮含量。就果园类型而言, 芸香科的效应值平均达0.418 (置信区间为0.332~0.505), 高于蔷薇科的0.166 (置信区间为0.121~0.215), 鼠李科效应值最低, 仅为0.056 (置信区间为−0.072~0.184)。就生草种类而言, 杂草和豆科的效应值分别为0.145 (置信区间为0.036~0.255)和0.171 (置信区间为0.102~0.179), 高于禾本科的0.065 (置信区间为0.005~0.131)。就气候类型而言, 半湿润区和湿润区无明显差异, 分别为0.128 (置信区间为0.078~0.185)和0.113 (置信区间为0.032~0.195), 略低于干旱区的0.182(置信区间为0.102~0.261)。就海拔而言, 高海拔效应值为0.251(置信区间为0.039~0.467), 高于低海拔的0.140(置信区间为0.11~0.167)。

图8 不同条件下生草对果园土壤全氮的影响Fig.8 Effects of sod-culture on soil total N in orchard under different factors
3 讨论
生草对果园生态系统服务功能的影响起初体现在物理状况的改变, 整合分析结果表明生草能够显著增加土壤含水量(图5)和降低土壤温度(图6)。首先, 生草后草被群落能够吸收转化太阳辐射, 增加反照率, 遮阴效果明显, 进而降低地表温度, 并且能够降低地表风速, 减少土壤蒸发; 同时生草后土壤容重降低、孔隙度增加、水稳性团聚体含量升高, 土壤持水能力增强, 在降雨期间增加入渗减少地表径流, 进而增加土壤含水量。另有研究指出生草对渭北旱地苹果()园土壤温度具有升温时降温和降温时保温的双向动态调控作用, 降低土壤温度日较差, 增加同一土层温度的稳定性等作用。也有研究表明, 生草覆盖虽然在土壤表层存在与果树争水的问题, 但在土壤深层能够缓和土壤干燥化程度, 有利于生态系统的可持续维护。综上, 生草对果园土壤温度和水分的调控有利于果树根系生长以及对养分的吸收利用, 促进土壤微生物活性, 对物质能量的转化具有积极作用, 进而影响土壤固碳、果实生产等其他生态系统服务功能。
随着生草年限的增加, 其对果园理化和生物性状的影响逐渐增强, 整合分析研究表明生草对果园土壤有机碳(图7)和全氮(图8)影响整体上均为显著正效应。生草后枯落物等有机残体会源源不断输入到果园生态系统中, 不断在土壤中腐烂矿化。凋落物与根际沉积物大量积累为微生物提供了充足的底物, 因此, 土壤微生物的多样性、活性及群落结构和功能得以显著提升。活性较高的微生物群落作为“加工者”通过分泌胞外酶分解或转化大分子植物源碳, 向土壤输送植物源残体, 从而提升土壤有机碳含量。同时, 微生物以植物源有机碳为食, 通过同化作用将其合成为自身生物量, 并在其细胞生长、数量增殖和死亡残体生成的过程中向土壤输送微生物源有机碳, 以“贡献者”的角色调控有机碳固定速率和有机碳库动态变化。草被群落根系的伸展牵拉作用在改善土壤容重、饱和导水率、大孔隙度等土壤物理特性的同时, 其分泌物及腐殖质将细小的土壤颗粒黏结形成较大的团粒结构, 而土壤团聚体能够将有机碳包裹起来, 通过空间上的物理隔离保护土壤有机碳, 使其免受微生物的分解利用, 从而提升土壤固碳能力。并且生草在降温和降低地表风速上的调控作用使土壤有机质的矿化过程受到抑制, 以CO形式流失的碳相对减少。土壤有机质是土壤中养分的主要来源, 众多研究表明土壤中有机碳和全氮含量为协同关系; 并且豆科牧草可以通过根瘤菌固氮, 因此具有更强的固氮效应(图8)。
生草对土壤水分和温度等微气象环境的改善, 对土壤有机碳和养分的提升为果树的生长发育提供了有利条件, 提高了果树在单位时间内对光照、二氧化碳、水分的转化效率, 增加含碳化合物的积累, 从而使得树势增强, 枝长叶厚, 果形美观, 内部品质等方面都有显著提高, 有利于增加果实产量和品质(图2-图4)。
可持续集约经营是当代农业发展的主流, 传统的清耕模式事倍功半, 在需要投入大量人力物力的同时, 也破坏了土壤结构、减少了土壤有机质含量、水土流失严重。与清耕果园相比, 生草增加了土壤有机碳、土壤养分、土壤水分, 改善了农田小气候, 促进了果园生态和经济效益的协同, 具有提质增效的作用。当前, 我国果园土壤管理仍以清耕为主, 全面因地制宜实施果园生草, 对促进我国果园健康可持续发展具有重大意义。
需要注意的是, 鉴于分析样本的限制, 本研究分析生态系统服务功能异质性因素时仅选取了海拔、气候、生草种类和果园类型4个因素, 实际上生草种植密度、果园管理方式等均会影响生态系统服务功能, 后续研究中将继续收集相关文献, 进一步梳理相关因子, 做出更加合理的分析。
4 结论
为探明生草对果园生态系统服务功能的影响, 本研究对全球已发表相关数据进行整合分析, 共获得了1387组数据。研究发现, 与清耕果园相比, 生草显著增加了果实产量, 改善部分果实品质, 提高了土壤水分, 降低了土壤温度, 改善了果园的微气候, 提升了果园的供给、调节和支持服务水平, 为我国果园通过生草实现可持续集约化提供了有力的证据。果园生草已成功应用到多种果园中, 但是由于该模式目前还存在一些问题和人们对该问题的认识不足, 影响了生草在果园栽培中的大范围推广。
猜你喜欢
中国记者(2022年8期)2022-09-15
军事文摘(2022年14期)2022-08-26
农家致富顾问·上半月(2021年6期)2021-12-26
山西果树(2020年1期)2020-12-10
农民致富之友(2020年33期)2020-12-03
学苑创造·A版(2020年11期)2020-01-07
阅读(低年级)(2019年9期)2019-11-15
环球时报(2019-05-23)2019-05-23
娃娃画报(2018年9期)2018-12-19
儿童故事画报·智力大王(2015年7期)2016-01-27
