
沉水植物生态化学计量学特征的区域差异以及生态修复的影响
2022-07-27 09:38赵钰殷春雨高弋明陈新芳关保华李宽意
生态科学 2022年4期
赵钰, 殷春雨, 高弋明, 陈新芳, 关保华,*, 李宽意
沉水植物生态化学计量学特征的区域差异以及生态修复的影响
赵钰1,2, 殷春雨1, 高弋明1,2, 陈新芳3, 关保华1,2,*, 李宽意1,2
1. 中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室, 南京 210008 2. 中国科学院大学, 北京 100049 3. 河海大学水文水资源学院, 南京 210098
为深入了解沉水植物生态化学计量学特征在西南高原地区和长江中下游平原地区湖泊中的差异, 以及生态修复的影响, 于2010年—2016年的夏季(6—9月)调查了36个湖泊的沉水植物。结果表明: (1)高原湖泊沉水植物群落的碳(C)含量显著高于平原湖泊, 氮(N)含量显著低于平原湖泊, 磷(P)含量高于平原湖泊但差异不显著; 高原湖泊沉水植物群落的C/N和C/P的比值显著高于平原湖泊, N/P显著低于平原湖泊; 在平原湖泊和高原湖泊共有的金鱼藻、苦草、穗花狐尾藻、微齿眼子菜和伊乐藻中, 仅有伊乐藻的N含量和金鱼藻的P含量与群落的差异性不一致。(2)在长江中下游平原地区生态修复后的水域中, 沉水植物群落的C和P含量显著高于未修复区, N含量低于未修复区但差异不显著; 修复区沉水植物群落的C/P和N/P显著低于未修复区, C/N显著高于未修复区; 修复区和未修复区共有的金鱼藻、苦草、穗花狐尾藻和竹叶眼子菜的C和P含量都跟群落的差异性一致。以上结果说明, 在清澈的水体中, 沉水植物的C和P含量高而N含量低; 湖泊富营养化会导致沉水植物N含量升高。因此, 在湖泊生态修复过程中, 应注重提高水体的透明度, 这将会提高沉水植物中C和P的含量, 从而有利于促进沉水植被区成为湖泊的C和P库。
沉水植物; 生态化学计量学特征; 区域差异; 湖泊; 生态修复
0 前言
在生物生长进化过程中, 生物有机体的碳(C)、氮(N)、磷(P)等化学元素的含量及其比例会保持动态稳定[1–3]; 但同时, 当有机体受某些环境影响时, 其化学计量学特征也会随之发生波动[4–6]。因此, 有机体化学计量学特征的变化能在一定程度上反映环境的变化[7–8]。生态化学计量学从细胞、个体、群落以及生态系统等不同等级层次的视角, 为研究生物与环境之间的关系和生物进化过程提供了一个新的工具。
沉水植物作为浅水湖泊中重要的初级生产者, 是复杂食物链形成的基础, 可以为其它水生生物提供物质和能量, 同时还可以净化水体, 是水体生物多样性赖以维持的基础[9–10]。沉水植物的生态化学计量学特征会随种类不同而发生变化[11–14], 同时, 沉水植物的化学计量学特征也深受水下光照条件和营养物质浓度的影响[15–18], 群落或物种水平的化学计量学特征的变化可以直接或间接地反映湖泊环境的状况及变化趋势[19–20]。
我国幅员辽阔, 拥有着数量众多、分布广泛、类型多样和成因不同的湖泊。不同区域的湖泊在物理、化学和生物特征等方面存在着显著差异[21], 水环境的差异将引起沉水植物群落和物种水平上的生态化学计量特征的不同。我国湖泊透明度总体表现为“西高东低”, 西部高原湖区的透明度要远高于东部平原湖区。长江中下游地区的湖泊多为浅水湖泊, 稠密的人口和频繁的经济活动导致大部分湖泊处于富营养化状态, 水体营养物质浓度普遍偏高且透明度较低[22]; 西南高原地区受人类活动影响较小, 光照强, 大部分湖泊相对较深, 水体富营养化程度尚浅, 透明度普遍较高[23–24], 两区域水环境的差异将导致沉水植物的生态化学计量学特征的不同。此外, 在富营养化湖泊中, 进行生态修复后, 水环境的改变也会引起沉水植物生态化学计量特征发生改变。
目前国内许多对沉水植物生态化学计量学特征的研究主要是集中在物种水平上的研究, 对群落水平上的研究还相对缺乏, 群落水平上沉水植物生态化学计量学特征的差异反应了其对不同环境条件适应的结果。其结果对于研究沉水植物的生长发育、植物多样性、生态系统过程以及预测群落变化有着重要意义, 可以为沉水植物保护及其在富营养化湖泊中的恢复提供依据。为此, 本研究比较了西南高原湖泊(青藏高原湖区和云贵高原湖区)与长江中下游平原湖泊(东部平原湖区)以及平原湖泊中富营养化湖泊和生态修复后湖泊中的沉水植物群落及物种水平上的生态化学计量学特征的差异, 以期深入了解水环境变化对沉水植物化学计量学特征的影响。
1 材料与方法
1.1 研究区概况
本研究数据来自2010年—2016年夏季(6—9月)的湖泊沉水植物采样调查(图1)。其中, 共调查长江中下游平原湖泊19个: 太湖、五里湖、高邮湖、溱湖、菜子湖、白荡湖、白马湖、升金湖、赤湖、团湖、严东湖、青凌湖、汤逊湖、梁子湖、豹澥湖、西凉湖、洪湖、洪泽湖和宝应湖; 西南高原湖泊17个: 抚仙湖、滇池、星云湖、洱海、阳宗海、拉市海、海西海、剑湖、天才湖、泸沽湖、程海、杞麓湖、巴松错、木地错、错鄂、时补错和越恰错。
在平原湖泊中, 五里湖与溱湖的局部区域进行过生态修复。五里湖是太湖伸入无锡的内湖, 五里湖生态修复区位于五里湖西北部一片名为渔夫湾的水域, 修复区面积约50000 m2, 平均水深2 m, 修复前透明度0.3 m, 年平均总磷(TP)浓度0.06 mg·L–1。溱湖修复区面积约80000 m2, 平均水深1.5 m, 修复前透明度0.2 m, 年平均TP浓度0.06 mg·L–1。五里湖水域在2010年6月至7月间进行生态修复, 溱湖水域在2011年5月至8月间进行生态修复, 修复措施均为清淤、除鱼和种植沉水植物。生态修复1个月后, 五里湖和溱湖TP浓度分别下降到0.02 mg·L–1和0.03 mg·L–1, 透明度上升到1.2 m和1.3 m。

图1 采样湖泊分布图
Figure 1 Distribution of sampling lakes
1.2 样品采集与处理方法
在每个湖泊的岸边到湖心设置3个样点, 每个样点采集1—2次沉水植物。使用采草夹将样点的沉水植物连根拔起, 用自来水清洗干净, 分类后选取每种植物的完整植株1—3株(总鲜重≥20 g), 在阴凉处风干后装入封口袋中, 置于冰箱中带回实验室; 在实验室中, 将采集回来的植物样品冷冻干燥至恒重, 并用粉碎机将植物样品粉碎后测定样品的C、N、P含量。其中, C和N含量采用元素分析仪(意大利EuroVector公司, EA3000)测定, P含量采用Prodigy电感耦合等离子体质谱仪(美国Leman公司, 7700x)测定。每个样点使用采水器在水下0.5 m处采集水样后装入500 mL采样瓶中。水样TP采用过硫酸钾消解、钼酸铵分光光度法测定, 水样TN采用过硫酸钾消解, 设备为紫外分光光度计(UV1750)。
1.3 数据处理
所得数据使用SPSS及R处理并作图[25], 均值比较采用独立样本T检验, 方差分析满足正态性和方差齐性的前提, 相关系数为Pearson相关系数。
2 结果与分析
2.1 研究区概况
在高原和平原湖泊中共采集到15种沉水植物(表1)。其中, 穿叶眼子菜和光叶眼子菜仅在高原湖泊中采集到, 大茨藻和水毛茛仅在平原湖泊中采集到, 其余种类的沉水植物在平原湖泊和高原湖泊皆采集到。
西南高原湖泊和平原湖泊水柱中TP浓度平均值分别为0.03 mg·L–1和0.04 mg·L–1; TN浓度平均值分别为1.18 mg·L–1和1.25 mg·L–1。长江中下游平原地区生态修复湖泊(五里湖和溱湖局部)和未修复湖泊(太湖、高邮湖、菜子湖、白荡湖、白马湖、升金湖、赤湖、团湖、严东湖、青凌湖、汤逊湖、梁子湖、豹澥湖、西凉湖、洪湖、洪泽湖、宝应湖)水柱中TP浓度平均值分别为0.02 mg·L–1和0.05 mg·L–1; TN浓度平均值分别为0.95 mg·L–1和1.32 mg·L–1。
2.2 高原湖泊和平原湖泊中沉水植物群落及物种水平的生态化学计量学特征差异
高原湖泊沉水植物的C含量显著高于平原湖泊的(0.05), N含量显著低于平原湖泊的(0.05), P含量高于平原湖泊但差异不显著(0.05)。

表1 采样湖泊中出现的沉水植物种类
高原湖泊沉水植物的C/N和C/P显著高于平原湖泊的(0.05), N/P显著低于平原湖泊的(0.05)。高原湖泊和平原湖泊沉水植物C/N的平均值分别为35.76和15.75; C/P的平均值分别为122.36和106.15; N/P的平均值分别为5.49和6.91。高原湖泊沉水植物中三种元素的比例约为196 C:5 N:1 P, 平原湖泊中约为109 C:7 N:1 P。
高原湖泊和平原湖泊中共有的沉水植物一共11种(表1), 我们对其中样本量超过10个的5种(金鱼藻、苦草、穗花狐尾藻、微齿眼子菜和伊乐藻)进行分析的结果显示: 这5种沉水植物C含量的平均值在高原湖泊中均大于其在平原湖泊中的C含量; 除伊乐藻外, 其余沉水植物N含量的平均值均小于其在平原湖泊中的N含量; 除金鱼藻外, 其余沉水植物P含量的平均值均大于其在平原湖泊中的P含量(图3)。
2.3 生态修复湖泊和未修复湖泊中沉水植物群落及物种水平的生态化学计量学特征差异
生态修复湖泊水域中沉水植物的C和P含量显著高于未修复区的(0.05; 图4), N含量低于未修复区的但无显著性差异(0.05)。
生态修复湖泊沉水植物的C/N显著高于未修复区的(0.05), C/P和N/P显著低于未修复区的(0.05)。生态修复湖泊和未修复湖泊中沉水植物C/N的平均值分别为17.29和15.50; C/P的平均值分别为100.37和130.05; N/P的平均值分别为5.94和8.39。生态修复湖泊中沉水植物的三种元素的比例约为103 C:6 N:1 P, 未修复湖泊中的约为130 C:8 N:1 P。
对生态修复湖泊和未修复湖泊中共有的4种沉水植物(金鱼藻、苦草、穗花狐尾藻和竹叶眼子菜) 进行分析的结果显示, 生态修复湖泊中4种沉水植物C和P含量的平均值均大于未修复湖泊(图5)。
3 讨论
本研究表明, 高原地区湖泊沉水植物的C含量(平均值375.70 mg·g–1)显著高于平原地区湖泊(平均值344.57 mg·g–1), 这一结果与李威等[26]对高原湖泊抚仙湖沉水植物的研究一致。段洪涛等[33]人通过GEE云计算平台对2000—2019年中国湖泊(20 km2)的透明度研究结果显示, 云贵高原湖区、青藏高原湖区和长江中下游平原湖区的透明度平均值分别为4.05 m、1.82 m和0.93 m, 可见西南高原湖泊的透明度要远大于长江中下游平原湖泊。沉水植物在透明度高的湖泊中光合作用强, 合成的碳水化合物多[27-30]; 另一方面, 在低光和高营养条件下, 沉水植物会积累游离氨基酸(FAA), 这个过程需要消耗大量碳水化合物[31-32]。这些原因可能共同导致了高原地区湖泊沉水植物C含量高于平原地区湖泊。此外, 尽管湖泊中无机碳的浓度高于空气, 但由于水中气体交换速率低, 尤其是水生植物叶面附近, 水会呈现出粘滞的状态, 严重影响CO2气体交换。因此, 生产力高的湖泊中, 无机碳常常被消耗贻尽, 成为限制初级生产者光合作用速率和生长的因素, 因而沉水植物会表现出生态化学计量特征上的低C含量[34–35]。高原湖泊大多处于贫营养到中营养的水平, 生产力相对低于平原富营养湖泊, 无机碳的制约作用较弱[36]。因此, 高原湖泊的沉水植物的C含量显著高于平原湖泊。
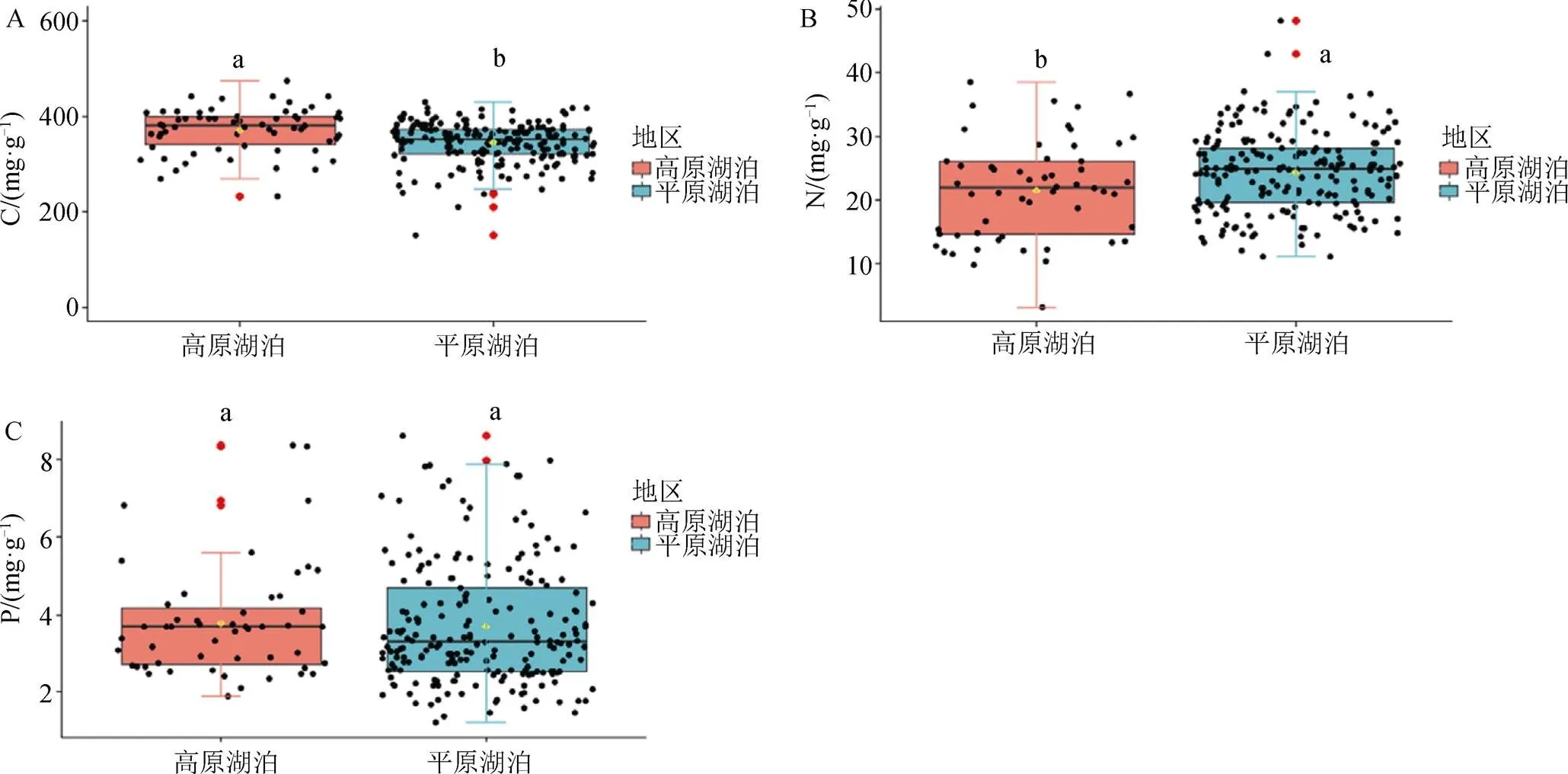
图2 西南高原地区和长江中下游平原地区湖泊中的沉水植物群落生态化学计量学特征(红点代表离群值, 黄点代表平均值)
Figure 2 Eco-stoichiometric characteristics of submerged macrophytes in lakes of southwest plateau area and plain area of the middle and lower reaches of the Yangtze River (The red points are outliers, and the yellow points are averages)

图3 不同种类沉水植物在西南高原湖泊与长江中下游平原湖泊中的生态化学计量学特征比较(P:平原湖泊; G:高原湖泊; J:金鱼藻; K:苦草; H:穗花狐尾藻; W:微齿眼子菜; Y:伊乐藻)
Figure 3 Eco-stoichiometric characteristics of submerged macrophytes among different species in lakes of the southwest plateau and the middle and lower reaches of the Yangtze River(P:plain lakes; G:plateau lakes; J:; K:; H:; W:; Y:)
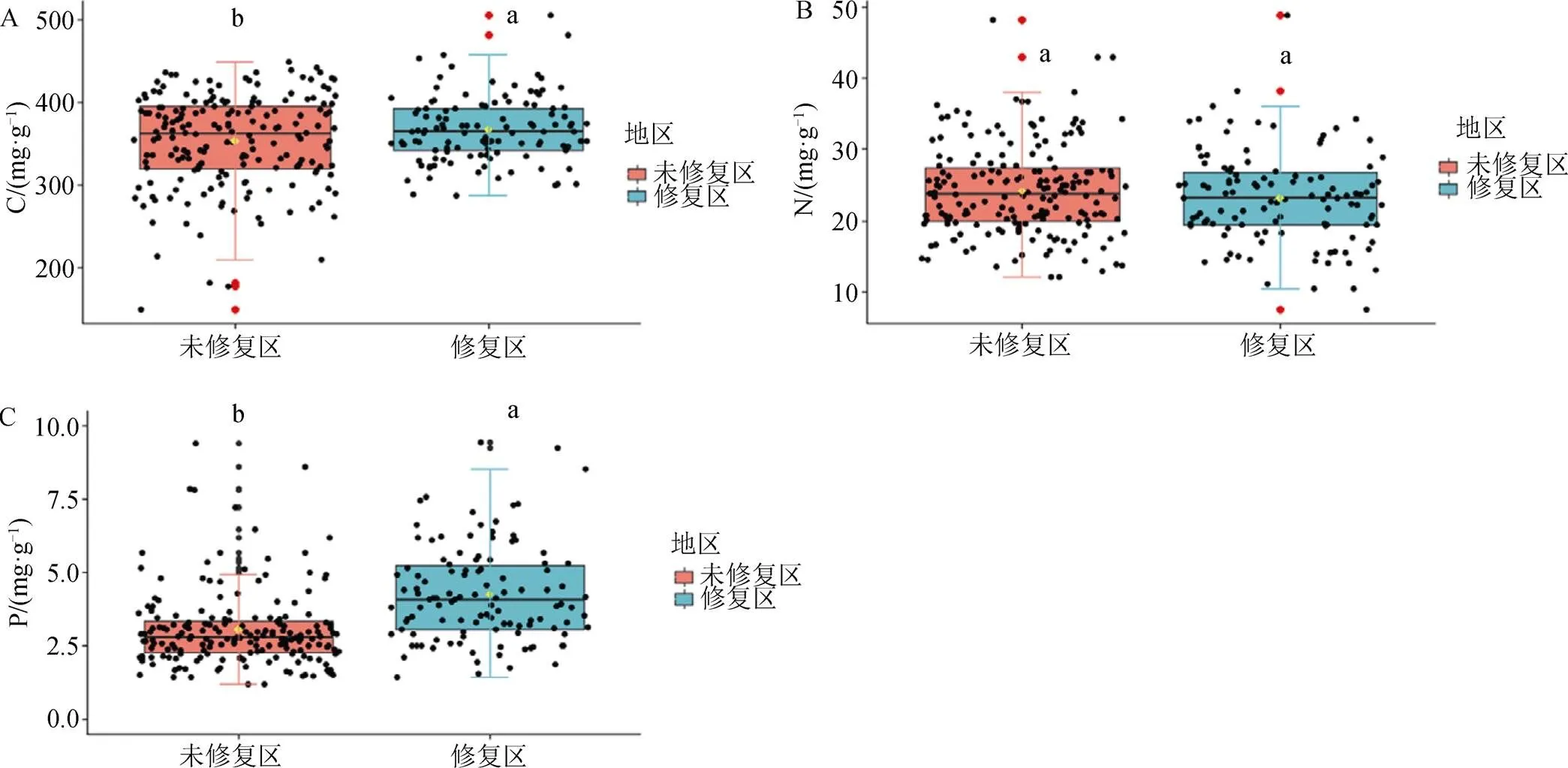
图4 长江中下游平原湖泊中生态修复区和未修复区中沉水植物群落的生态化学计量学特征
Figure 4 Eco-stoichiometric characteristics of submerged macrophytes inthe ecological restored lakes and the unrestored lakes

图5 不同种类沉水植物在长江中下游生态修复湖泊水域和未修复水域的生态化学计量学特征(R:生态修复区; U:未修复区; J:金鱼藻; K:苦草; H:穗花狐尾藻; M:竹叶眼子菜)
Figure 5 Eco-stoichiometric characteristics of different submerged macrophytes between the ecological restored lakes and the unrestored lakes(R:ecological restoration region; U:unrestored region; J:; K:; H:; M:)
湖泊中营养盐的含量决定沉水植物N和P含量[37]。长江中下游平原地区湖泊的氮负荷高于高原地区湖泊; 平原地区人类活动频繁, 雨量充沛, 大气中N的干、湿沉降量都大于高原地区[38]。因此, 平原地区沉水植物可吸收利用的N远远超过高原地区。此外, 在低光高营养条件下, 植物体内会过度积累NH4+-N[39–41]。平原地区湖泊多呈现富营养化状态, 沉水植物体内的高NH4+-N也可能导致了总N含量高于高原地区湖泊的沉水植物。
高原地区湖泊中水柱的P含量普遍低于平原地区湖泊, 沉水植物可获得的磷应该是低于平原地区湖泊的。但结果却显示, 高原地区湖泊中沉水植物的P含量反倒是高于平原地区的。这可能是不同地区的沉水植物的生长速率不同导致的。根据生长速率假说, 生物个体的生长速率主要体现在P含量的变化上, 高生长速率的生物个体的P含量也高[42]。高原地区夏季短暂, 沉水植物的生长季节短, 平均生长速率较高, 这可能是高原地区湖泊中的沉水植物P含量平均值要高于平原地区的原因之一。这一结果也支持温度—植物生理假说, 该假说认为来自较冷和高海拔生境的植物表现出较高的叶片养分浓度, 因为温度可以直接影响生理过程如光合作用和呼吸作用的速率[43]。因此, 高原地区湖泊沉水植物较高的P含量可能是由于植物对低温和寒冷生境的适应和生理驯化, 从而增强了酶的活性并加快了新陈代谢和生长速度, 使得其P含量高。此外, 高的光可利用性也会增加沉水植物对P的吸收[44], 高原地区湖泊中光的可利用性高, 因此其沉水植物的P含量高于平原地区。但是高原湖泊沉水植物P含量与平原地区湖泊差异并不显著, 这说明沉水植物P含量的弹性较大, 其受到环境因素的影响也更复杂。
此外, 由于不同物种的化学计量学存在差异, 因此, 群落物种组成的差异也将会对沉水植物群落的化学计量学特征产生重要影响[45]。本研究从高原湖泊和平原湖泊共获取15种沉水植物, 其中有11种沉水植物为两个区域湖泊中共有。平原地区修复湖泊和未修复湖泊共获取13种, 其中有10种沉水植物为两类湖泊中共有。群落物种的高度相似降低了物种组成差异带来的影响, 因此, 不同区域沉水植物群落的差异更多地体现了水环境的差异。环境差异和群落组成差异对群落化学计量学特征差异的贡献率需要进一步研究。
长江中下游平原地区生态修复湖泊的透明度和营养盐状况与高原湖泊相似: 透明度高、水柱中P含量低。因此, 在生态修复后的水域中, 沉水植物群落的C和P含量表现出与高原湖泊—平原湖泊相似的差异, 即生态修复湖泊中沉水植物的C和P含量高于透明度低且营养丰富的未修复区, 而N含量低于未修复区。
4 结论
结果表明, 无论是高原地区湖泊还是平原地区生态修复后的湖泊, 沉水植物群落的生态化学计量学特征都表现出C和P含量高而N含量低的趋势; 透明度可能是解释沉水植物的化学计量学特征的重要因素。本研究的结论是: 在湖泊生态修复过程中, 提高水体的透明度能提高沉水植物中C和P的含量, 从而有利于促进沉水植被区成为湖泊的C和P库。
[1] ÅGREN G I. The C:N:P stoichiometry of autotrophs theory and observations[J]. Ecology Letters, 2004, 7(3): 185–191.
[2] ELSER J J, DOBBERFUHL D R, MACKAY N A, et al. Organism size, life history, and N:P stoichiometry[J]. Bioscience, 1996, 46(9): 674–684.
[3] ZHANG Lixia, BAI Yongfei, HAN Xingguo. Application of N:P stoichiometry to ecology studies[J]. Acta Botanica Brasilica, 2003, 45(9): 1009–1018.
[4] ANDERSEN T, ELSER J J, HESSEN D O. Stoichiometry and population dynamics[J]. Ecology Letters, 2004, 7(9): 884–900.
[5] ELSER J J. Ecological stoichiometry: from sea lake to land[J]. Trends in Ecology & Evolution, 2000, 15(10): 393– 394.
[6] ELSER J J, FAGAN W F, KERKHOFF A J, et al. Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change[J]. New Phytologist, 2010, 186(3): 593–608.
[7] ELSER J J, STERNER R W, GOROKHOVA E, et al. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2000, 3(6): 540–550.
[8] 邢伟, 吴昊平, 史俏, 等. 生态化学计量学理论的应用、完善与扩展[J]. 生态科学, 2015, 34(1): 190–197.
[9] BORNETTE G, PUIJALON S. Macrophytes: Ecology of Aquatic Plants[M]. Holand: Academic Press, Elsevier, 2009.
[10] DE KLUIJVER A, NING J, LIU Z, et al. Macrophytes and periphyton carbon subsidies to bacterioplankton and zooplankton in a shallow eutrophic lake in tropical China[J]. Limnology and Oceanography, 2015, 60(2): 375–385.
[11] BORNETTE G, PUIJALON S. Response of aquatic plants to abiotic factors: a review[J]. Aquatic Sciences, 2011, 73(1): 1–14.
[12] LI Wei, CAO Te, NI Leyi, et al. Size-dependent C, N and P stoichiometry of three submersed macrophytes along water depth gradients[J]. Environmental Earth Sciences, 2015, 74(5): 3733–3738.
[13] WANG Zhong, XIA Chengxing, YU Dan, et al. Low- temperature induced leaf elements accumulation in aquatic macrophytes across Tibetan Plateau[J]. Ecological Engineering, 2015, 75(6): 1–8.
[14] ZHANG Peiyu, KURAMAE A, VAN LEEUWEN C H A, et al. Interactive Effects of rising temperature and nutrient enrichment on aquatic plant growth, stoichiometry, and palatability[J]. Frontiers in Plant Science, 2020, 11(58): 1–15.
[15] GONZALEZ SAGRARIO M A, JEPPESEN E, GOMA J, et al. Does high nitrogen loading prevent clear-water conditions in shallow lakes at moderately high phosphorus concentrations?[J]. Freshwater Biology, 2005, 50(1): 27– 41.
[16] SU Haojie, WU Yao, XIE Ping, et al. Effects of taxonomy, sediment, and water column on C:N:P stoichiometry of submerged macrophytes in Yangtze floodplain shallow lakes, China[J]. Environmental Science and Pollution Research, 2016, 23(22): 57–85.
[17] CHAMBERS P A. Light and nutrients in the control of aquatic plant community structure. 2. Insitu observations[J]. Journal of Ecology, 1987, 75(3): 621–628.
[18] VERMAAT J E, DEBRUYNE R J. Factors limiting the distribution of submerged waterplants in the lowland river vecht (The Netherlands)[J]. Freshwater Biology, 1993, 30(1): 147–157.
[19] SU Haojie, WU Yao, XIA Wulai, et al. Stoichiometric mechanisms of regime shifts in freshwater ecosystem[J]. Water Research, 2019(149): 302–310.
[20] CHOI J Y, JEONG K S, KIM S K, et al. Role of macrophytes as microhabitats for zooplankton community in lentic freshwater ecosystems of South Korea[J]. Ecological Informatics, 2014(24): 177–185.
[21] 曹金玲, 许其功, 席北斗, 等. 我国湖泊富营养化效应区域差异性分析[J]. 环境科学, 2012, 33(6): 1777–1783.
[22] 黄亮, 吴莹, 张经, 等. 长江中游若干湖泊水生植物体内C, N, P及δ^13C分布[J]. 地球学报, 2003, 24(6): 515– 518.
[23] SHEN Ming, DUAN Hongtao, CAO Zhigang, et al. Sentinel-3 OLCI observations of water clarity in large lakes in eastern China: Implications for SDG 6.3.2 evaluation[J]. Remote Sensing of Environment, 2020(247): 111950.
[24] 郝贝贝, 吴昊平, 史俏, 等. 云南高原10个湖泊沉水植物的碳、氮、磷化学计量学特征[J]. 湖泊科学, 2013, 25(4): 539–544.
[25] REESE R A. Graphics for statistics and data analysis with R, 2nd edition[J]. Journal of The Royal Statistical Society Series A-Statistics In Society, 2018, 181(4): 1261–1262.
[26] 李威, 符辉, 曹特, 等. 抚仙湖沉水植物分布及其碳、氮和磷化学计量学特征[J]. 湖泊科学, 2017, 29(2): 448– 457.
[27] Sterner R W, Elser J J, Fee E J, et al. The light: nutrient ratio in lakes: the balance of energy and materials affects ecosystem structure and process[J]. American Naturalist, 1997, 150(6): 663–684.
[28] Sterner R W, Elser J J. Ecological stoichiometry: The biology of elements from molecules to the biosphere[M]. Princeton: Princeton University Press, 2002.
[29] ZHANG Yunlin, ZHOU Yongqiang, SHI Kun, et al. Optical properties and composition changes in chromophoric dissolved organic matter along trophic gradients: Implications for monitoring and assessing lake eutrophication[J]. Water Research, 2018(131): 255–263.
[30] HESSEN D O, AGREN G I, ANDERSON T R, et al. Carbon, sequestration in ecosystems: The role of stoichiometry[J]. Ecology, 2004, 85(5): 1179–1192.
[31] ZHANG Meng, CAO Te, NI Leyi, et al. Carbon, nitrogen and antioxidant enzyme responses ofto both low light and high nutrient stresses[J]. Environmental and Experimental Botany, 2010, 68(1): 44–50.
[32] XING Wei, WU Haoping, HAO Beibie, et al. Stoichiometric characteristics and responses of submerged macrophytes to eutrophication in lakes along the middle and lower reaches of the Yangtze River[J]. Ecological Engineering, 2013(54): 16–21.
[33] LIU Dong, DUAN Hongtao, LOISELLE S, et al. Observations of water transparency in China's lakes from space[J]. International Journal of Applied Earth Observation and Geoinformation, 2020(92): 102187.
[34] MABERLY S C. Diel, episodic and seasonal changes in pH and concentrations of inorganic carbon in a productive lake[J]. Freshwater Biology, 1996, 35(3): 579–598.
[35] YAN X, YU D, LI Y K. The effects of elevated CO2on clonal growth and nutrient content of submerge plant Vallisneria spinulosa[J]. Chemosphere, 2006, 62(4): 595– 601.
[36] KRAGH T, SAND-JENSEN K. Carbon limitation of lake productivity[J]. Proceedings of the Royal Society B-Biological sciences, 2018, 285(1891): 20181415.
[37] AZCóN R, AMBROSANO E, CHAREST C. Nutrient acquisition in mycorrhizal lettuce plants under different phosphorus and nitrogen concentration[J]. Plant Science, 2003, 165(5): 1137–1145.
[38] COLLINS K A, LAWRENCE T J, STANDER E K, et al. Opportunities and challenges for managing nitrogen in urban stormwater: A review and synthesis[J]. Ecological Engineering, 2010, 36(11): 1507–1519.
[39] VON WIREN N, GAZZARRINI S, GOJON A, et al. The molecular physiology of ammonium uptake and retrieval[J]. Current Opinion in Plant Biology, 2000, 3(3): 254–261.
[40] CRONIN G, LODGE D M. Effects of light and nutrient availability on the growth, allocation, carbon/nitrogen balance, phenolic chemistry, and resistance to herbivory of two freshwater macrophytes[J]. Oecologia, 2003, 137(1): 32–41.
[41] REICH P B, OLEKSYN J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001–11006.
[42] ELSER J J, BRACKEN M E S, CLELAND E E, et al. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems[J]. Ecology Letters, 2007, 10(12): 1135–1142.
[43] WEIH M, KARLSSON P S. Growth response of Mountain birch to air and soil temperature: is increasing leaf-nitrogen content an acclimation to lower air temperature?[J]. New Phytologist, 2001, 150(1): 147–155.
[44] ZHANG Meng, CAO Te, NI Leyi, et al. Light-dependent phosphate uptake of a submersed macrophyte[J]. Aquatic Botany, 2011, 94(4): 151– 157.
[45] Xia Chengxing, Yu Dan, Wang Zhong, et al. Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China[J]. Ecological Engineering, 2014(70): 406–413.
Regional differences and the effects of ecological restorationon stoichiometric characteristics of submerged macrophytes
ZHAO Yu1,2, YIN Chunyu1, GAO Yiming1,2, CHEN Xinfang3, GUAN Baohua1,2,*, LI Kuanyi1,2
1. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing210008, China 2. University of Chinese Academy of Sciences, Beijing100049,China 3. Hydrology and Water Resources College, Hohai University, Nanjing210098, China
Submerged macrophytes were surveyed in 36 lakes from June to September during 2010-2016 across the southwest plateau and the east plain of the middle and lower reaches of the Yangtze River. The study aimed to determinethe ecological stoichiometric characteristics of submerged macrophytes between the plateau and plain lakes and to know the impact of restoration on eco-stoichiometric characteristics of submerged macrophytes in plain lakes. The results showed that the carbon (C) content of submerged macrophytes community in plateau lakes was significantly higher than in plain lakes but the nitrogen (N) content was significantly lower than in plain lakes. The phosphorus (P) content was higher in plateau lakes than in plain lakes, but not significant. The C/N and C/P in submerged macrophytes communities were significantly higher in plateau lakes than in plain lakes, while the N/P was the opposite. Among the five submerged macrophytes that emerged in the two regions (,,,and), only the N content of theandthe P content of thewere not consistent with the trend of submerged macrophytes community. In the plain lakes, the C and P contents of submerged macrophytes of restored lakes were significantly higher than unrestored lakes. The C/P and N/P in restored lakes were significantly lower than that in the unrestored lakes, while the C/N was the opposite. Among the four submerged macrophytes that emerged in both restored and unrestored lakes, the trends in C and P contents were consistent with the submerged macrophytes community. The above results implied that submerged macrophytes had high C and P contents but low N content in clear-water lakes. Eutrophication might lead to high N content in macrophytes. Therefore, improving the transparency in the lake restoration is beneficial to turn submerged macrophytes region into the reservoirs of C and P for lakes.
submerged macrophytes; eco-stoichiometric characteristics; regional differences; lakes; ecological restoration
10.14108/j.cnki.1008-8873.2022.04.003
Q178.1
A
1008-8873(2022)04-016-09
2020-08-03;
2020-10-11
浙江省‘尖兵’‘领雁’研发攻关计划(2022C02038); 国家自然科学基金(32171534)
赵钰(1995—), 男, 山东泰安人, 硕士研究生, 主要从事湖泊生态学研究, E-mail: zhaoyu181@mails.ucas.ac.cn
通信作者:关保华, 女, 博士, 副研究员, 主要从事湖泊生态学研究, E-mail: bhguan@niglas.ac.cn
赵钰, 殷春雨, 高弋明, 等. 沉水植物生态化学计量学特征的区域差异以及生态修复的影响[J]. 生态科学, 2022, 41(4): 16–24.
ZHAO Yu, YIN Chunyu, GAO Yiming, et al. Regional differences and the effects of ecological restoration on stoichiometric characteristics of submerged macrophytes[J]. Ecological Science, 2022, 41(4): 16–24.
猜你喜欢
黄河之声(2022年6期)2022-08-26
华人时刊(2022年3期)2022-04-26
世界科学技术-中医药现代化(2021年5期)2021-11-05
湖泊科学(2021年4期)2021-07-07
世界科学技术-中医药现代化(2021年12期)2021-04-19
环境科学导刊(2020年6期)2020-12-07
安徽文学(2020年10期)2020-10-26
花卉(2020年12期)2020-01-09
——以武汉紫阳湖为例
绿色科技(2019年2期)2019-05-21
滇池(2019年1期)2019-02-14
