
马铃薯HSP90 基因家族的全基因组鉴定及高温干旱胁迫下的表达分析
2022-07-15 06:27芳乔岩王永强李茜李丹王亚士柴薇薇
西北农业学报 2022年6期
杨 芳乔 岩王永强李 茜李 丹王亚士柴薇薇
(1.陇东学院 农林科技学院,甘肃庆阳 745000;2.陇东旱地作物种质改良及产业化协同创新中心,甘肃庆阳 745000;3.甘肃省陇东生物资源保护利用与生态修复重点实验室,甘肃庆阳 745000;4.甘肃农业大学 农学院,兰州 730000;5.西北民族大学 生命科学与工程学院,兰州 730000)
马铃薯(Solanum tuberosumL.)是全世界最重要的粮食作物之一,具有较高的投入产出比,其适应性强、耐寒、耐旱、耐瘠薄,但不耐高温,高温导致马铃薯茎节间伸长、叶片变小,降低光合效率,块茎发生退化、畸形,生长停滞,造成严重减产[1]。近百年来,中国地表平均气温升高幅度约为0.5~0.8 ℃,增温速率为0.08 ℃/10 a,是全球气候变暖最显著的国家之一;近50 a来的温度增幅尤为明显,西北地区的增温速率为0.33℃/10 a,明显高于全国平均水平[2]。极端高温事件给西北地区马铃薯的生长发育和生产安全带来了巨大挑战,因此提高马铃薯非生物胁迫抗性具有重要意义。作物在长期进化过程中形成了适应不良环境的分子机制,在面对高温等逆境胁迫时,为避免细胞内稳态受到破坏,细胞内会迅速合成大量的分子伴侣蛋白,这类蛋白被称为热激蛋白(HSP,Heat shock protein)。HSP蛋白具有重要调控功能,有助于错误折叠蛋白质恢复正常构象并维持细胞稳态,保护植物在高温、寒冷、干旱、盐等逆境胁迫下生存[3]。在植物中,HSP 可分为5个家族:HSP60、HSP70、HSP90、HSP100和小分子热激蛋白(s HSP,Small heat shock protein)家族,其中HSP90是进化上高度保守,分布较广的一类热激蛋白,在细胞质、叶绿体、线粒体、内质网内均有分布,占细胞总蛋白的1%~2%,当遭遇胁迫时含量能提高4%~6%[3-4]。
HSP90蛋白在栽培番茄(Solanum lycopersicumL.)、马铃薯、辣椒(Capsicum annuumL.)等作物的耐热性、抗旱性等研究中得到验证[5-7]。高温胁迫后[8]:(1)HSP90 能结合折叠和变性的蛋白质,维持细胞内蛋白质的稳态;(2)HSP90能激活底物蛋白质的活性,进而发挥其生理功能;(3)HSP90 能抑制热激因子(HSF,Heat shock factor)的功能,负向调控包含热激元件(HSE,Heat shock element)基因的转录。研究发现,过表 达At HSP90.2、At HSP90.5、At HSP90.7会降低拟南芥(Arabidopsis thaliana)对盐胁迫和干旱胁迫的耐性,但会提高其对高钙离子的耐性,而At HSP90.2在ABA 依赖型信号通路中发挥重要作用[9]。Yamada 等[10]发现,拟南芥At HSP90.2负调控热诱导基因的转录,热激后At HSP90.2暂时失活,进而激活HSF 以提高热诱导基因的表达。Gil等[11]证实,拟南芥HSP90.1可激活底物蛋白ZTL,进而减少不溶性蛋白质聚集、重新折叠错误蛋白,提高植物的耐热性。拟南芥HSP90.1 能 与ROF1 和NBR1 互 作,通 过 热应激记忆提高其耐热性[12-13]。Wang 等[14]发现HSP90能激活SGT1b-TIR1蛋白复合体,通过生长素信号转导控制植物在高温胁迫下的生长发育。Samakovli等[15-16]发现,HSP90在YODA 级联介导的热应激反应中发挥重要功能,调控气孔的形成以适应高温环境。过表达Os HSP50.2可提高水稻(Oryza sativaL.)对干旱、渗透胁迫的抗性,在干旱胁迫下转化株的超氧化物歧化酶(Superoxide dismutase,SOD)活性、脯氨酸含量则显著提高[17]。Xu等[18]发现,在拟南芥过表达5个Gm HSP90s基因均可增加脯氨酸含量,提高其高温、干旱等非生物胁迫抗性。Wei等[19]发现,木薯(Manihot esculenta)Me HSP90.9能通过招募MeWRKY20和MeCatalase1蛋白调控木薯的抗旱性。深入研究植物HSP90基因在逆境胁迫下的功能,对于正确认识植物胁迫信号转导、多重胁迫响应机制及提高作物的抗逆性具有重要的应用价值。目前,HSP90基因家族在作物中的非生物胁迫响应机制、表达模式及其提高作物抗逆性的机制尚不明晰,其中马铃薯HSP90基因家族的相关研究较少。本研究以茄科植物HSP90基因家族为研究对象,对马铃薯全基因组数据进行结构域搜索,鉴定HSP90基因家族成员,明确HSP90基因家族在基因组中的分布特征和启动子的顺式作用元件,并使用qRT-PCR 技术分析其高温与干旱胁迫下的表达差异;构建HSP90基因家族蛋白质相互作用网络(Protein-protein interaction,PPI),对HSP90伙伴蛋白的功能进行解析,为研究马铃薯HSP90基因家族调控高温干旱等逆境胁迫的重要功能提供依据。
1 材料与方法
1.1 试验材料的培养与处理方法
试验材料选择对高温干旱有较强耐性的马铃薯野生种恰克薯(Solanum chacoenseBitter),材料由陇东学院农林科技学院提供,组培苗培养条件为温度22~24 ℃,光照时长12 h/d;继代培养使用的是普通MS培养基。试验设2个处理(高温胁迫处理,干旱胁迫处理),3 次重复,对照(CK)为常温下生长的组培苗。高温胁迫处理:将25 ℃生长5周的组培苗置入40 ℃培养箱中处理1 h,6 h。干旱胁迫处理:PEG 模拟水分胁迫参照邓珍等[20]的方法,将15% PEG(PEG-8000)胁迫下生长的组培苗置入25℃生长3周;取出不同处理下的叶片,液氮速冻后-76 ℃超低温冰箱保存。
1.2 茄科植物HSP90 基因家族的鉴定
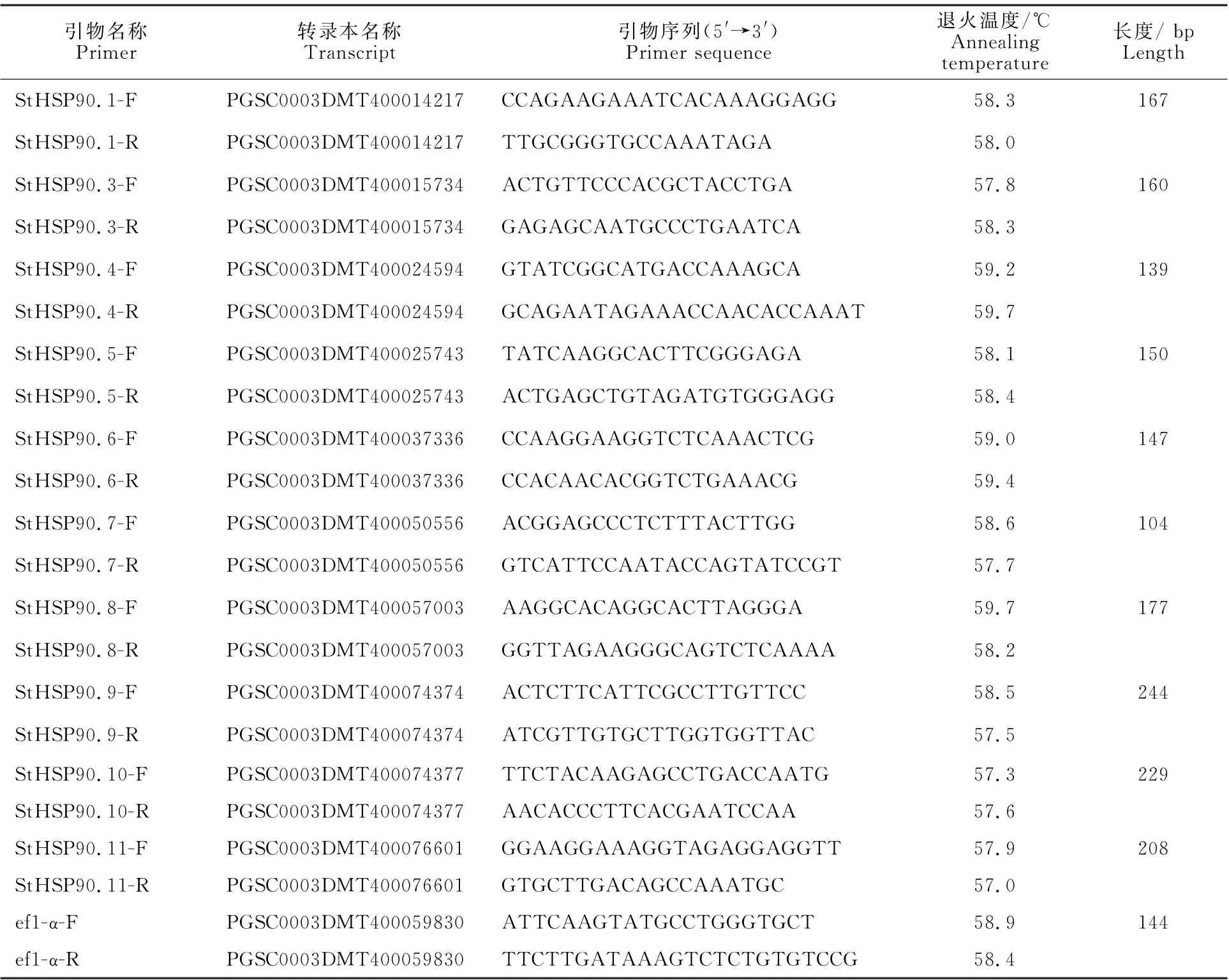
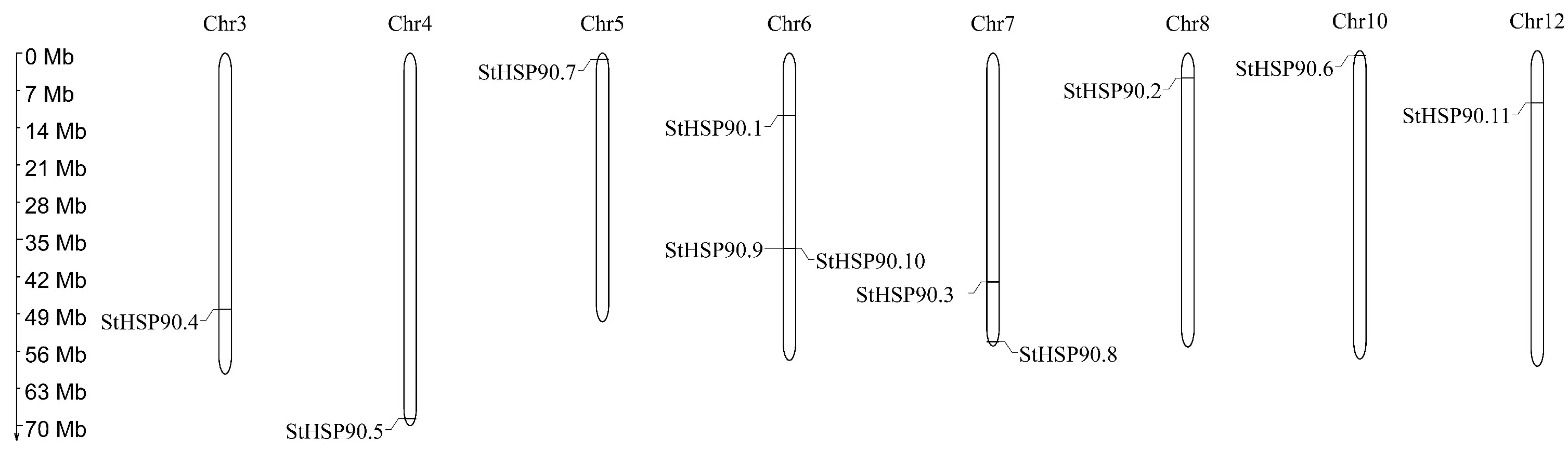
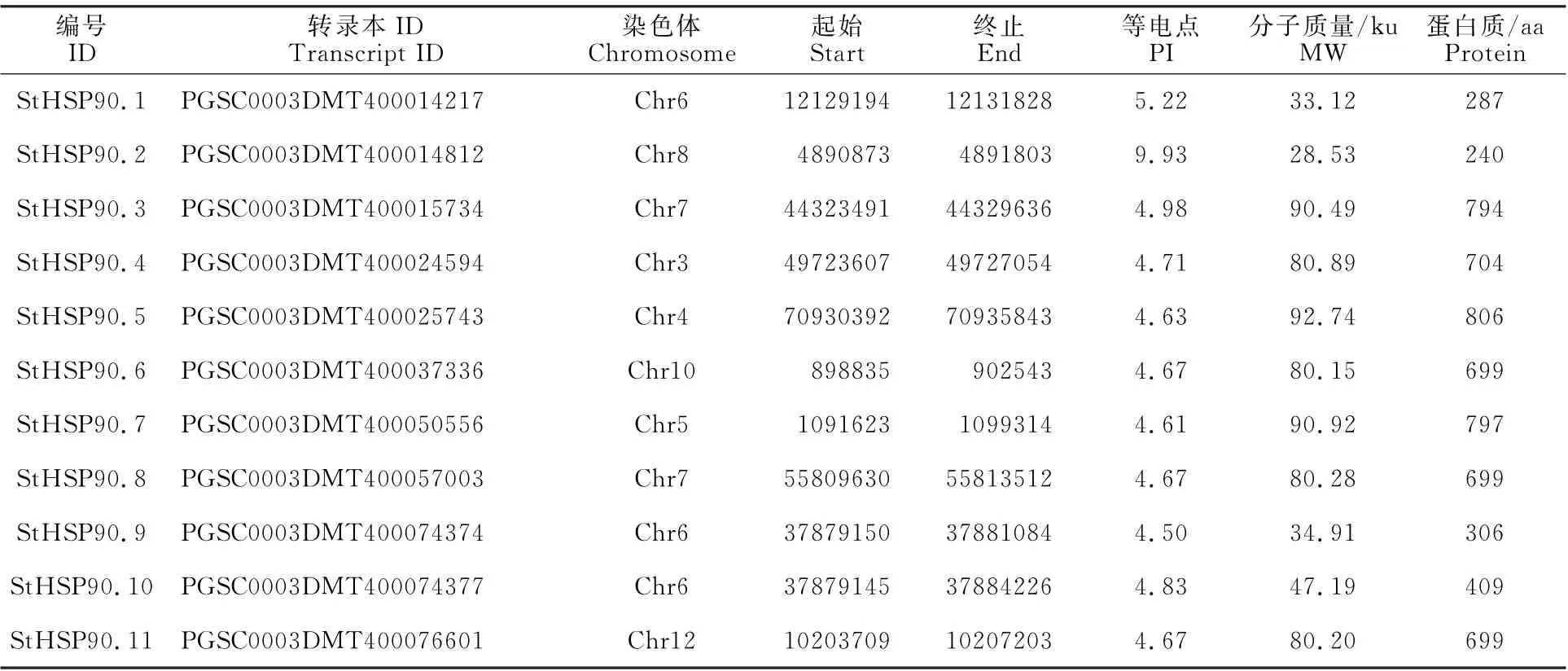
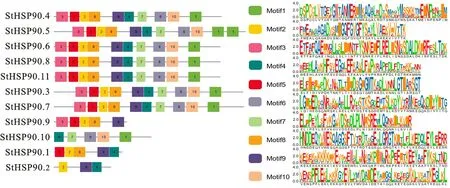
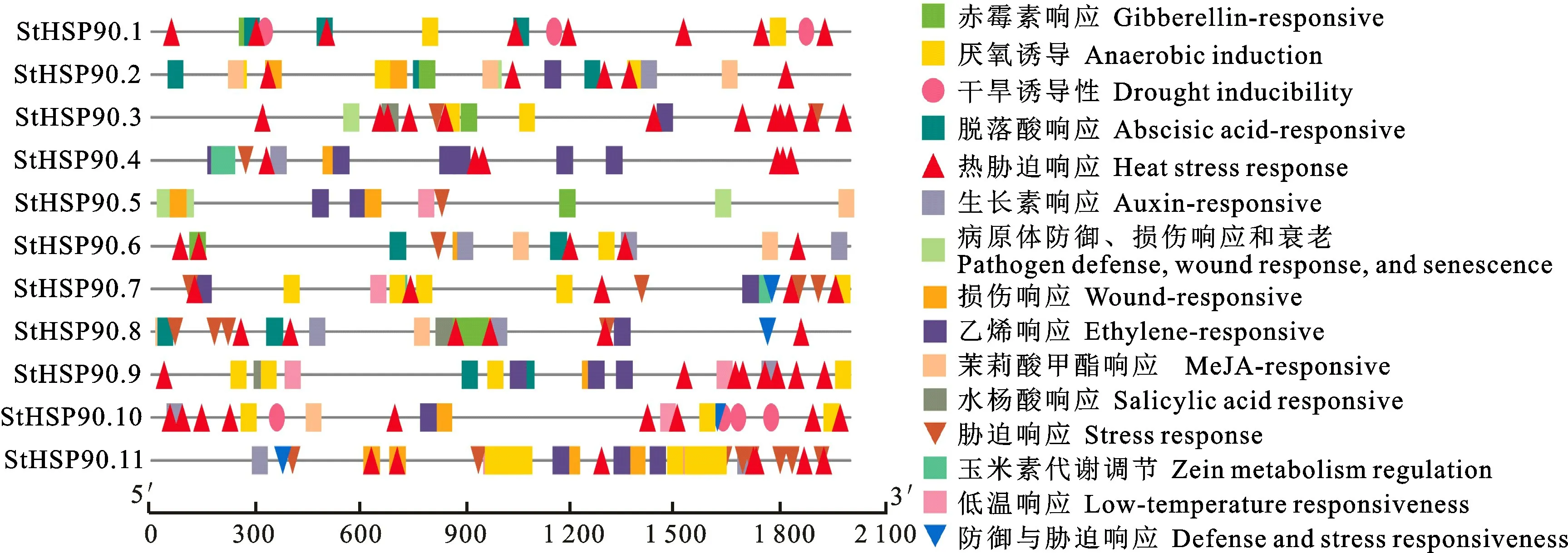
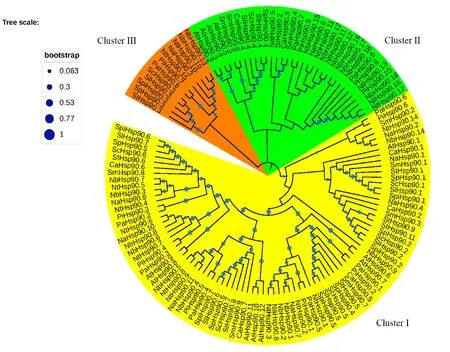
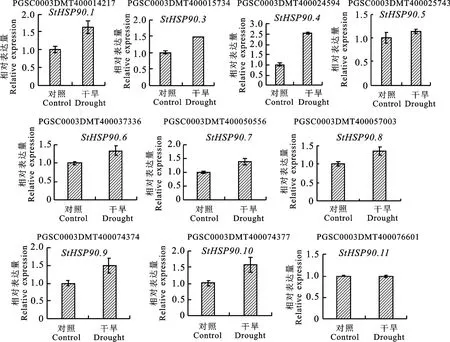
在Pfam26.0(http://pfam.xfam.org/)下载HSP90蛋白(ID:PF00183)的HMM 文件。在茄科植物基因组网站(https://solgenomics.net/)下载12个茄科植物蛋白质序列:栽培番茄、马铃薯、渐窄叶烟草(Nicotiana attenuatatorr.Ex.Watson)、辣椒、红花烟草(Nicotiana tabacumL.)、本氏烟草(Nicotiana benthamiana)、腋花矮牵牛(Petunia axillaris)、矮牵牛(Petunia inflata)、潘那利番茄(Solanum pennellii)、醋栗番茄(Solanum pimpinellifolium)、智利番茄(Solanum chilense)、栽培茄(Solanum melongena),同时下载拟南芥HSP90蛋白质序列。利用本地化HMMER(http://www.hmmer.org/)子程序hmmsearch从茄科植物及拟南芥蛋白质中筛选出HSP90蛋白ID(E-value 根据马铃薯基因注释文件,利用MG2C(http://mg2c.iask.in/mg2c_v2.0/)进行马铃薯HSP90基因家族的染色体定位。使用MEME Suite 5.1.1(http://meme-suite.org/)分析马铃薯HSP90蛋白的保守基序,最大保守基序数设为10,其余参数为默认设置。用MEGA-X 构建茄科植物HSP90家族的系统进化树(最大似然法,bootstrap value为1 000)。使用TBtools软件对马铃薯HSP90基因家族成员在干旱胁迫和热胁迫下的基因表达量进行分析[21]。 利用PLANTCARE 软件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析HSP90 家族基因启动子顺式作用元件;使用Zhao等[22]提供的python程序分析马铃薯HSP90家族基因启动子的热激元件HSE,使用TBtools对启动子功能元件进行可视化[21]。 为研究与HSP90家族成员发生作用的底物蛋白,使用STRING(https://string-db.org/)预测与HSP90 家族发生相互作用的功能伙伴蛋白[23],选择score>0.9的蛋白质作为功能伙伴蛋白,使用Bingo对功能伙伴蛋白进行GO 富集分析(correctedP-value<0.05)。 采用UNIQ-10 柱式Trizol总RNA 抽提试剂盒(上海生工,B511321)提取马铃薯试管苗总RNA,用Maxima Reverse Transcriptase(Thermo Scientific,EP0743)进行反转录。将c DNA 样品稀释10倍作为模板上机检测,选择ef1-α作为内参基因[24],重复3次,实时定量PCR 试剂使用2X SG Fast qPCR Master Mix (High Rox,B639273,BBI),使用ABI Stepone plus型荧光定量PCR 仪(ABI,Foster,CA,USA)进行实时荧光检测,引物序列见表1,其中St HSP90.2(PGSC0003DMT400014812)基因表达量过低无法正常扩增。数据处理采用2-△△Ct计算样本处理前后的相对表达量,△△Ct=(△Ct处理样品-△Ctef1a)-(△Ct对照样品-△Ctef1a)。 表1 荧光定量PCR 引物序列Table 1 Primers sequences for qRT-PCR 经过hmmsearch 程序和Pfam 数据库的筛选,去冗余后得到11 个马铃薯HSP90基因家族成员,对其编号(St HSP90.1~St HSP90.11);基于马铃薯基因组注释文件,使用MG2C 软件对HSP90家族成员进行染色体定位(图1)。 图1 马铃薯HSP90 基因家族成员染色体定位Fig.1 Chromosome location of family members of potato HSP90 genes 根据遗传连锁图可知,马铃薯St HSP90基因家族成员分布于8 条染色体上。Chr3、Chr4、Chr5、Chr8、Chr10、Chr12 染色体上各分布1 个HSP90基因家族成员,Chr7 染色体分布2 个HSP90基因家族成员,Chr6 染色体分布3 个HSP90基因家族成员;HSP90基因家族的等电点为4.50~9.93,分子质量为28.53~92.74 ku(表2)。 表2 马铃薯HSP90 基因家族成员Table 2 Family members of potato HSP90 genes 在11个St HSP90基因中,XPASY ScanProsite(https://prosite.expasy.org/scanprosite/)分析发现11个St HSP90基因家族成员中有8个具有PS00298 序列(Heat shock hsp90 proteins family signature),而St HSP90.1、St HSP90.2、St HSP90.10中不具有该序列。利用MEME 工具对HSP90基因家族的保守基序分析,共含有10个保守基序(图2)。其中7个HSP90家族成员(St HSP90.3、St HSP90.4、St HSP90.5、St HSP90.6、St HSP90.7、St HSP90.8、St HSP90.11)含有完整的5 个保守基序。7 个成员中,St HSP90.4、St HSP90.6、St HSP90.8 和St HP90.11的结构相似,St HSP90.3 和St HSP90.5 结构相似,表明这些蛋白质具有类似功能。11个HSP90家族成员均包含HSP90典型结构域(PF00183),9个HSP90家族成员(St HSP90.1、St HSP90.2、St HSP90.3、St HSP90.4、St HSP90.5、St HSP90.6、St HSP90.8、St HSP90.9、St HSP90.11)同时包含2个特征结构域(HSP90 和HATPase_c 结构域)。 图2 马铃薯HSP90 基因家族蛋白保守基序Fig.2 Family protein conserved motifs of potato HSP90 genes 对马铃薯HSP90基因上游序列分析后发现,HSP90基因普遍存在与热胁迫相关的HSE 元件(图3),HSE 元 件 从5~12 个 不 等;其 中St HSP90.3含 有12 个HSE 元 件,而St HSP90.5不包含HSE 元件。其次,9个基因含有1~8个与干旱响应与胁迫响应元件,其中St HSP90.11含有8 个STRE 元 件(stress response element),St HSP90.1有3 个干旱响应元件,St HSP90.10含有4个干旱响应元件。5个基因共包含7个低温响应元件,其中St HSP90.3含有2个低温响应元件(图3)。此外,HSP90基因中包含有大量与赤霉素、脱落酸、茉莉酸、玉米素、乙烯等激素响应元件(图3);激素响应元件中乙烯响应元件最多,St HSP90.2~St HSP90.11共9 个基因包含22个乙烯响应元件,其中St HSP90.4包含6个乙烯响应元件;其次为茉莉酸应答元件,St HSP90.2、St HSP90.3、St HSP90.6、St HSP90.8、St HSP90.10共5个基因包含18 个茉莉酸应答元件,其中St HSP90.2和St HSP90.8各包含6个茉莉酸应答元件(图3)。St HSP90.1、St HSP90.2、St HSP90.6、St HSP90.8、St HSP90.9共包含14 个脱落酸响应元件,其中St HSP90.1包含4 个脱落酸响应元件。部分HSP90基因包含11 个生长素响应元件,8个赤霉素响应元件,4个玉米素响应元件和3个水杨酸响应元件(图3)。 图3 马铃薯HSP90 基因顺式作用元件Fig.3 Cis-regulatory elements of potato HSP90 genes STRING 数 据 库 分 析 显 示11 个HSP90 蛋白构成复杂的相互作用网络(图4-A,interaction score>0.700),该网络有21个节点和130条边,平均节点度为12.4,PPI富集P-value 图4 马铃薯HSP90 基因家族蛋白质互作网络分析Fig.4 Integrative analysis of HSP90 protein-protein interaction(PPI)network in potato 利用STRING 数据库预测到HSP90基因家族的22个功能伙伴蛋白(表3,score≥0.9),对这22个蛋白质进行GO 功能富集分析,发现其生物学过程富集可分为6大类(图4-B):(1)蛋白质的折叠、水解与稳定性调节相关途径;(2)氧水平应答、热激应答于热适应等高温胁迫响应途径;(3)胁迫应答、盐胁迫应答、渗透胁迫应答与非生物刺激等非生物胁迫途径;(4)内质网核信号转导、细胞内信号转导等信号转导通路;(5)花分生组织决定性、去黄化、营养期转变和花粉管生长等生长发育途径;(6)生物刺激应答、真菌刺激应答与真菌防御反应等生物胁迫途径。22个功能伙伴蛋白在KEGG 数据库共注释到植物病原菌互作(sot04626),内质网蛋白加工(sot04141),蛋白质输出(sot03060)和吞噬小体(sot04145)4 个通路。 表3 利用STRING 数据库预测的与HSP90伙伴蛋白Table 3 Predicted functional partners of HSP90 using STRING database 利用hmmsearch从12个茄科植物中共鉴定出130个HSP90基因家族成员,构建系统发育树(图5),系统树显示HSP90基因家族被分为三簇(Ⅰ、Ⅱ、Ⅲ),簇Ⅰ中有85个基因家族成员,簇Ⅱ中有33个基因家族成员,簇Ⅲ中有12个基因家族成员。簇Ⅰ中包含12个茄科植物的HSP90基因家族成员,其中的马铃薯HSP90家族成员与潘那利番茄、智利番茄、栽培番茄、醋栗番茄等茄科番茄属植物的HSP90基因具有较高同源性,亲缘关系较近(图5)。簇Ⅱ中包含11个茄科植物成员(除潘那利番茄)的33 个基因家族成员,包括St HSP90.2,St HSP90.4。马铃薯St HSP90.2与辣椒Ca HSP90.3、栽培茄Sm HSP90.4和渐窄叶烟草Na HSP90.5具有较近的亲缘关系;马铃薯St HSP90.4与智利番茄Sc HSP90.4栽培番茄Sl-HP90.4和醋栗番茄Spi HSP90.4具有较近的亲缘关系。St HSP90.2和St HSP90.4与红花烟草、本氏烟草、矮牵牛、腋花矮牵牛的亲缘关系较远。簇Ⅲ包括9个茄科植物(马铃薯、潘那利番茄、醋栗番、栽培番茄、智利番茄、矮牵牛、红花烟草、本氏烟草、渐窄叶烟草)的12个基因家族成员,其中马铃薯St HSP90.3与智利番茄Sc HSP90.3,栽培番茄Sl HP90.3,醋栗番茄Spi HSP90.3和潘那利番茄Sp HSP90.3的同源性较高,但是与渐窄叶烟草、红花烟草、本氏烟草的亲缘关系较远。 图5 茄科植物HSP90 基因家族系统进化树Fig.5 Phylogenetic tree of HSP90 gene family in Solanaceae plants 采用qRT-PCR 技术对马铃薯HSP90基因家族成员在高温、干旱胁迫下的表达量进行分析。结果表明,高温胁迫后,10个马铃薯HSP90基因的表达量均呈上调趋势(图6)。仅St HSP90.2(PGSC0003DMT400014812)基因的表达量过低无法正常扩增。6个基因呈现出先升高后降低的趋势,其中4 个基因的表达量极高:St HSP90.9(PGSC0003DMT400074374)在胁迫1 h后上调至4 190倍,6 h后降至1 621倍;St HSP90.10(PGSC00003DMT400074377)上调2 800 倍,在6 h后降低至1 429 倍;St HSP90.1(PGSC00003 DMT400014217)上调2 139倍,6 h后降至1 126倍;St HSP90.4(PGSC0003DMT 400024594)上调1 090 倍,6 h 后 降 至468 倍。St HSP90.3(PGSC0003DMT400015734),St HSP90.5(PGSC 0003DMT400025743),St HSP90.7(PGSC0003DMT400050556)和St HSP90.8(PGSC0003DMT400057003)4个基因在高温胁迫1~6 h时呈现出持续上调的表达趋势;只有St HSP90.8(PGSC0003DMT400057003)仅上调1.5 倍。在干旱胁迫3 周后,与对照相比仅St HSP90.11(PGSC0003DMT400076601)表达量略有下降,其余9个马铃薯HSP90基因的表达量呈上调趋势,上调幅度为1.1~2.5倍,其表达量明显小于高温胁迫下的基因表达量(图7)。其中St HSP90.4(PGSC0003DMT400024594)表达量最高,上调2.5倍,St HSP90.1(PGSC0003DMT400014217)和St HSP90.10(PGSC0003DMT400074377)上调1.6倍,St HSP90.9(PGSC0003DMT400074374)上调1.5倍,其余5个HSP90基因上调幅度均小于1.5倍。 图6 马铃薯HSP90 基因家族成员基因在高温胁迫下的qRT-PCR 分析Fig.6 q RT-PCR analysis of HSP90 in potato under high temperature stress 图7 马薯HSP90 基因家族成员基因在干旱胁迫下的qRT-PCR 分析Fig.7 qRT-PCR analysis of HSP90 in potato under drought stress HSP90基因家族在进化过程中高度保守,在遭受非生物胁迫时能被激活,进而调节植物的新陈代谢和细胞内环境稳态。本研究从茄科植物中鉴定出130个HSP90基因,在马铃薯基因组中鉴定出11个HSP90基因,其在染色体上的分布规律与番茄、辣椒HSP90相似[5,7]。系统进化分析显示HSP90基因进化上高度保守,12 个茄科植物的130个HSP90基因分为3簇,与水稻和辣椒相同;马铃薯HSP90基因家族与潘那利番茄、智利番茄、栽培番茄等番茄属植物亲缘关系较近。7个马铃薯HSP90家族成员具有10个蛋白保守基序,且位置相对一致;所有HSP90家族成员均包含PF00183 结 构 域,9 个HSP90 家 族 成 员 包 含HSP90和HATPase_c结构域。 植物启动子顺式作用元件决定基因的转录起始点和转录效率,能调控下游基因的表达。热激因子HSF能感受多途径转导的热信号,识别并结合HSE 元件[25],通过热激启动子(Heat shock promoter)促进热胁迫基因表达[26]。本研究发现马铃薯HSP90基因家族普遍存在HSE元件,位置和数量分布存在差异;St HSP90.5未发现HSE元件,热激后St HSP90.5仅上调1.5倍,而其他HSP90基因热胁迫后发生不同程度的上调,这与HSE 元件分布的位置及结构不同相关;Zhao等[22]发现,不同类型和结构的HSE 元件,序列变异和位点错配的HSE 元件均会影响下游基因转录效率,这与本研究结果一致。除HSE元件之外,马铃薯HSP90基因存在干旱响应元件、STRE 元件、低温响应元件等胁迫响应元件,表明马铃薯HSP90基因参与多重胁迫抗性调控。研究表明[27-28],植物HSP 蛋白/伙伴蛋白受干旱胁迫、激素信号诱导,且乙烯信号通路也是植物响应干旱胁迫的重要途径。本研究发现,长期干旱胁迫后多数马铃薯HSP90基因明显上调,其中St HSP90.4上调2.5倍;除St HSP90.1外其余马铃薯HSP90基因均包含乙烯响应元件,而St HSP90.4包含6 个乙烯响应元件,这表明马铃薯HSP90基因通过乙烯信号通路参与干旱胁迫调控。本研究发现马铃薯HSP90基因非编码区包含生长素、茉莉酸、脱落酸、乙烯等激素响应顺式元件,受相应激素信号途径调控;这与前人研究结果类似,植物HSP90 蛋白通过生长素信号途径[29]、茉 莉 酸 信 号 途 径[30]、ABA 信 号 转 导 通路[31]与乙烯信号通路[32-33]调控逆境胁迫。综上所述,马铃薯HSP90基因启动子受多种激素信号调控,预示马铃薯HSP90基因通过激素信号转导通路应对逆境胁迫。 HSP90蛋白能通过信号转导、激酶和转录因子等蛋白质调控植物生理生化过程,能与HSP70及其他蛋白发生作用,进而调控蛋白质的折叠[34]。本研究证实BIP 蛋白属于HSP70家族,是马铃薯HSP90基因家族PPI网络的中心蛋白。研究表明[35-36],BIP 蛋白在内质网质量控制中发挥重要作用,在未折叠蛋白反应(unfolded protein response,UPR)中促进内质网蛋白质折叠,缓解非生物胁迫引起的内质网损伤,过表达BIP基因的大豆表现出较强抗旱性。本研究中,高温与干旱胁迫后多数马铃薯HSP90基因明显上调,马铃薯HSP90 家族蛋白均与BIP 蛋白发生互作,高温干旱胁迫下高表达基因St HSP90.5与BIP蛋白有极强的互作关系(score>0.9),说明该HSP90-BIP复合体能调控UPR 反应,在抵抗极端逆境引起的内质网胁迫中发挥重要作用。目前,植物中已发现上百种蛋白质受HSP90蛋白质调控,通过HSP90伙伴蛋白质复合体参与逆境胁迫应答、生长发育、信号转导等多种生物学过程[8,29,37]。马 铃 薯St HSP9.6、St HSP90.8和St HSP9.11在高温或干旱胁迫下表达量较高,这3个HSP90基因的伙伴蛋白CHIP(PGSC0003-DMT400051983)与拟南芥E3 泛素连接酶At-CHIP高度同源;Zhou等[38]发现AtCHIP蛋白通过泛素-蛋白酶体系统降解高温等胁迫产生的错误折叠蛋白,这表明马铃薯HSP90蛋白调控毒蛋白降解通路,在高温等逆境胁迫具有重要功能。Pant等[39]发现在高温胁迫下,拟南芥CPN60ATPXL1蛋白复合体通过激活NADP-苹果酸脱氢酶活性抑制ROS形成,进而提高其耐热性和抗病性;而马铃薯St HSP90.7伙伴蛋白CPN60A(PGSC0003DMT400002933)与拟南芥CPN60A 蛋白高度同源,且St HSP9.7在高温与干旱胁迫后表达量较高,说明St HSP90.7在高温或干旱胁迫过程中发挥重要调控作用。 本研究发现马铃薯HSP90蛋白与其伙伴蛋白构成一个复杂的逆境胁迫网络,鉴定的HSP90-伙伴蛋白复合体在高温、干旱等逆境胁迫下发挥重要作用,但需要进一步使用酵母双杂交技术、双分子荧光互补技术对 HSP90-BIP、HSP90-CHIP与HSP90-CPN60A 蛋白质之间的互作关系进行充分验证,从而为提高马铃薯的多重胁迫抗性和培育抗旱耐热型品种提供新途径。1.3 马铃薯HSP90 基因家族特征分析
1.4 HSP90 基因家族的基因结构与启动子分析
1.5 HSP90家族蛋白质互作分析
1.6 荧光实时定量PCR分析
2 结果与分析
2.1 马铃薯HSP90 基因家族成员鉴定与蛋白质基序分析
2.2 HSP90 基因家族成员的顺式作用元件分析
2.3 HSP90 基因家族成员PPI网络与功能伙伴蛋白注释分析


2.4 茄科植物HSP90 基因家族系统进化分析
2.5 马铃薯HSP90 基因家族成员的qRT-PCR分析

3 讨论
猜你喜欢
传感器世界(2022年6期)2022-11-25今日农业(2022年16期)2022-11-09今日农业(2022年15期)2022-09-20环球时报(2022-06-20)2022-06-20中国核电(2021年3期)2021-08-13作文大王·笑话大王(2018年12期)2018-03-23中学生数理化·中考版(2016年7期)2016-12-07中学物理·高中(2016年8期)2016-08-08创新作文(5-6年级)(2015年9期)2015-10-21红蜻蜓(2015年4期)2015-06-01
