
盐碱胁迫对甘蓝型油菜生理及分子机制的影响
2022-07-11 08:07:50杨明煊刘志文
华北农学报 2022年3期
李 班,吕 莹,杨明煊,宋 婷,于 放,刘志文
(大连工业大学 生物工程学院,辽宁 大连 116034)
土壤盐碱化作为世界各国正在面临着的重大环境问题,对于农作物的产量和质量有着严重的影响,据不完全统计,全世界盐碱地的面积为9.5亿hm2,其中我国为9 913万hm2,而且土壤盐碱化趋势也正逐年增加[1-2]。盐碱胁迫包括盐胁迫、碱胁迫以及盐碱共同胁迫,其中,盐胁迫主要是由中性盐NaCl和Na2SO4引起,而碱胁迫则是由碱性盐NaHCO3和Na2CO3引起[3-7]。盐碱胁迫对植物带来的伤害包括渗透胁迫、离子毒害、活性氧胁迫等[8]。在受到盐碱胁迫的时候,植物一般会通过2种途径来调节自身渗透平衡,一种是植物细胞会通过吸收外界环境中的Na+、K+、Cl-等无机盐离子来调节;另一种是植物细胞通过自身合成的脯氨酸、可溶性糖、甜菜碱、抗氧化酶等物质来调节[9];而甜菜碱作为植物体内无毒的渗透调节物质,它的合成以胆碱为底物,经过2步催化形成,一是胆碱加氧酶(CMO),催化胆碱氧化成甜菜碱醛;二是甜菜碱醛脱氢酶(BADH),催化甜菜碱醛形成甜菜碱[10]。在植物受到盐碱胁迫时,叶绿素含量变化可以反映光合作用变化以及植物对盐碱胁迫的响应[11]。另外,在受到盐碱胁迫时,植物体内会产生一种有毒物质—丙二醛,丙二醛的过量积累会影响植物细胞的正常生理活动,因此,丙二醛可以作为衡量植物受到盐碱胁迫强弱的指标之一[12]。
甘蓝型油菜(BrassicanapusL.)作为我国重要油料作物,具有长势强、产量高、营养丰富、经济价值高、耐盐碱能力强等诸多优点[13]。此外,土壤上层所积累的大量沉积性有机物及土壤中的部分难溶物,都会在甘蓝型油菜根系所分泌的有机酸的作用下加速溶解,这不仅能改善土壤,还能减轻地力消耗,兼有养地的作用,因此具有极大的种植潜力[14]。
一直以来,关于植物耐盐碱方面的报道多集中在单一盐、单一碱对植物胁迫后其生理生化指标方面的研究[15-17],关于盐碱共同胁迫及其分子机制的研究较少,并且在较多研究中,盐碱胁迫被统称为盐胁迫,缺少一定的科学性。实际上,在我国大面积的盐碱环境中,植物是受到了盐、碱共同胁迫的影响[18],因此,盐碱共同胁迫是目前生产实际中存在的主要问题。本试验主要利用人工模拟盐胁迫、碱胁迫以及盐碱胁迫处理甘蓝型油菜,从生理及分子机制两方面研究甘蓝型油菜对盐碱胁迫的响应,旨在为提高甘蓝型油菜耐盐碱能力以及增加甘蓝型油菜产量提供重要的理论依据。
1 材料和方法
1.1 试验材料
试验材料为甘蓝型油菜华油杂62,由华中农业大学提供。
1.2 试验方法
1.2.1 混合盐碱溶液的配制和处理 试验中所用复合盐成分为NaCl、Na2SO4,按照1∶1混合;复合碱成分为Na2CO3、NaHCO3,按照1∶1混合;复合盐碱成分为NaCl、Na2SO4、Na2CO3、NaHCO3,按照1∶1∶1∶1混合。共设置处理组3个(复合盐、复合碱、复合盐碱),对照组1个(ddH2O)。每个处理组设置3个梯度,其中复合盐包括:Y25(含盐量25 mmol/L)、Y50(含盐量50 mmol/L)、Y75(含盐量75 mmol/L);复合碱包括:J25(含碱量25 mmol/L)、J50(含碱量50 mmol/L)、J75(含碱量75 mmol/L);复合盐碱包括:YJ25(含盐碱量25 mmol/L)、YJ50(含盐碱量50 mmol/L)、YJ75(含盐碱量75 mmol/L)。
1.2.2 种子发芽率测定 将50粒籽粒饱满、大小一致的华杂62种子播种到含有不同人工模拟的复合盐、复合碱和复合盐碱溶液的平板上培养,萌发期间规律性喷洒相应溶液,以保证种子发芽期需要的水分,7 d 后计算发芽率,每个处理设3 次重复。
1.2.3 生理指标的测定 为减少试验误差,本试验中所用幼苗均由一颗华杂62种子经NaClO消毒后再经植物组织培养进行大量扩培,随后将生长30 d且长势一致的组培苗移入营养土中,待生长14 d后,取长势一致的油菜幼苗进行不同浓度的复合盐、复合碱和复合盐碱溶液处理,分别处理7,14,21 d后,收集叶片,迅速存于-80 ℃超低温冰箱中,用于不同生理指标的测定,每个处理设3次重复。
叶绿素含量测定采用分光光度法,脯氨酸含量测定采用酸性茚三酮法;可溶性糖含量测定采用蒽酮比色法,SOD采用氮蓝四唑(NBT)光还原法测定[19],丙二醛含量测定采用硫代巴比妥酸法,CAT活性采用过氧化氢法测定,POD活性采用愈创木酚法测定[20],甜菜碱含量测定方法参考文献[21]进行。
1.2.4 实时荧光定量分析 通过TRIzol法提取经不同条件处理后的甘蓝型油菜幼苗叶片中的总 RNA,利用反转录酶将所提取的RNA反转录成cDNA,以PP2A作为内参进行实时荧光定量分析。利用甘蓝型油菜中甜菜碱合成途径关键酶基因CMO的全序列,通过Primer Premier 5软件根据引物设计原则设计特异性引物,引物序列如表1所示。

表1 引物序列Tab.1 Primer sequences
实时荧光定量仪器为LightCycle®480,使用 SYBR GREEN Ⅰ荧光染料,程序如下:95 ℃预变性5 min;95 ℃变性20 s,50 ℃退火15 s,72 ℃延伸15 s,40 个循环;72 ℃延伸10 min。反应体系:SYBR GREEN Ⅰ 10 μL;Primer 1 0.2 μL;Primer 2 0.2 μL;cDNA 2 μL;ddH2O 7.6 μL,总体系20 μL。将得到的Ct值使用2-ΔΔCt方法计算,每个基因测3组平行,计算其平均数以及标准误差。
1.3 数据处理与分析
通过Microsoft Excel 2010软件进行数据的计算和整理,采用 SPSS 软件对数据进行方差分析和最小显著差异性检验,通过Origin 2018进行图表的绘制。
在所有图表中,误差线表示3个生物学重复的标准误差,不同的小写字母表示在P<0.05 水平上各试验处理间差异显著。
2 结果与分析
2.1 盐碱胁迫对甘蓝型油菜种子发芽率的影响
随着盐碱溶液浓度的升高,甘蓝型油菜的发芽率有所下降,当油菜种子受到中高浓度(YJ50、YJ75)盐碱胁迫影响时,发芽率为0,其中复合盐处理的种子发芽率与CK组相比变化差异较小,说明复合盐对油菜种子发芽率影响最小,其次为复合碱的伤害,而复合盐碱对种子发芽率影响程度最大,且达显著水平(P<0.05)(表2)。

表2 盐碱胁迫对种子发芽率的影响Tab.2 The effect of saline-alkali stress on the seed germination
2.2 甘蓝型油菜耐盐碱机制
2.2.1 叶绿素含量 如图1所示,在经低中复合盐(Y25、Y50)处理油菜幼苗时,叶绿素含量随处理时间延长先升高后降低,当处理时间为14 d,叶绿素的含量最高,分别为16.53,16.67 mg/g,在经过高复合盐(Y75)、复合碱、复合盐碱溶液处理后,叶绿素含量随着处理浓度增加与处理时间的延长而下降。因此,低浓度盐碱溶液会促进叶绿素的积累,而高浓度盐碱溶液会抑制叶绿素的积累,其中以复合碱与复合盐碱对叶绿素的影响最为显著(P<0.05)。
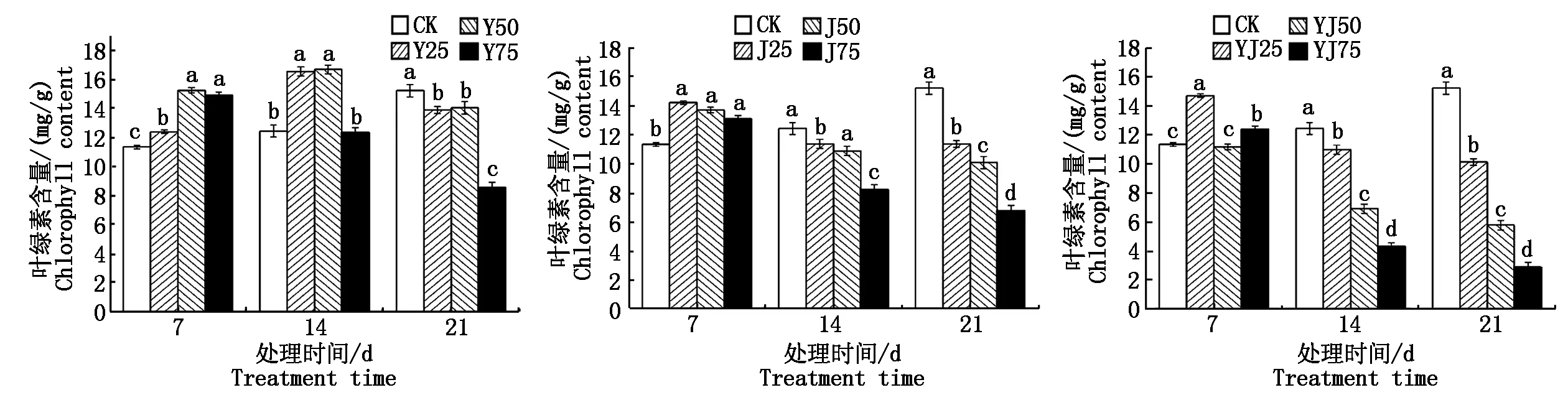
不同小写字母表示差异显著(P<0.05)。图2—11同。Different lowercase letters indicate significant difference(P<0.05).The same as Fig.2—11.
2.2.2 脯氨酸和可溶性糖含量 如图2,3所示,随着盐碱溶液浓度升高与处理天数的增加,叶片中脯氨酸及可溶性糖含量显著增加,当胁迫处理21 d,中高复合盐(Y50、Y75)与高复合盐碱(YJ75)处理组中脯氨酸含量急剧升高,分别是对照组的12.19,12.97,65.99倍,差异显著(P<0.05);高复合盐碱(YJ75)组中可溶性糖大量积累,胁迫21 d可溶性糖积累量高达40.60 mg/g,为对照组的5.21倍。

图2 复合盐、复合碱、复合盐碱对甘蓝型油菜叶片脯氨酸含量的影响Fig.2 Effect of compound salt,compound alkali,compound saline-alkalion the proline content in leaves of Brassica napus L.

图3 复合盐、复合碱、复合盐碱对甘蓝型油菜叶片可溶性糖含量的影响Fig.3 Effect of compound salt,compound alkali,compound saline-alkalion the soluble sugar content in leaves of Brassica napus L.
2.2.3 丙二醛含量 如图4所示,在复合盐、复合碱、复合盐碱溶液胁迫油菜7 d,其体内丙二醛含量变化差异较小,在中高复合盐(Y50、Y75)处理幼苗14,21 d,丙二醛含量要高于对照组,且差异显著;胁迫21 d,高复合碱(J75)处理油菜幼苗后的丙二醛含量要高于高复合盐(Y75)处理后的丙二醛含量;复合盐碱对油菜幼苗丙二醛含量的影响趋势与复合盐、复合碱的影响趋势相似。

图4 复合盐、复合碱、复合盐碱对甘蓝型油菜叶片丙二醛含量的影响Fig.4 Effect of compound salt,compound alkali,compound saline-alkalion malondialdehyde content in leaves of Brassica napus L.
2.2.4 抗氧化酶活性 由图5可知,复合盐处理可显著降低SOD活性;在复合盐处理7 d,POD的活性随着盐浓度的升高而升高,与空白对照相比,高盐(Y75)处理后的POD活性提高了3.03倍,且随处理时间不同,POD活性先升后降,在第14 天达到最高值;在复合盐处理7 d,处理组中CAT活性均高于对照组,差异显著(P<0.05),在胁迫14 d,CAT活性随着盐浓度呈先降后升趋势,且随处理时间的增加,CAT的活性降低。
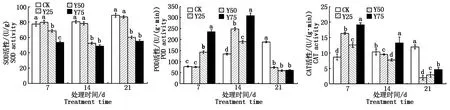
图5 盐胁迫对甘蓝型油菜叶片抗氧化酶活性的影响Fig.5 Effect of salt stress on antioxidant enzyme activity in leaves of Brassica napus L.
如图6所示,复合碱处理后,处理组中SOD活性均低于对照组;在胁迫14 d,复合碱处理后的POD活性达到最大值,分别为286.5,349.25,302.12 U/(g·min);复合碱处理后的CAT活性随处理时间延长与处理浓度增加而显著降低(P<0.05)。
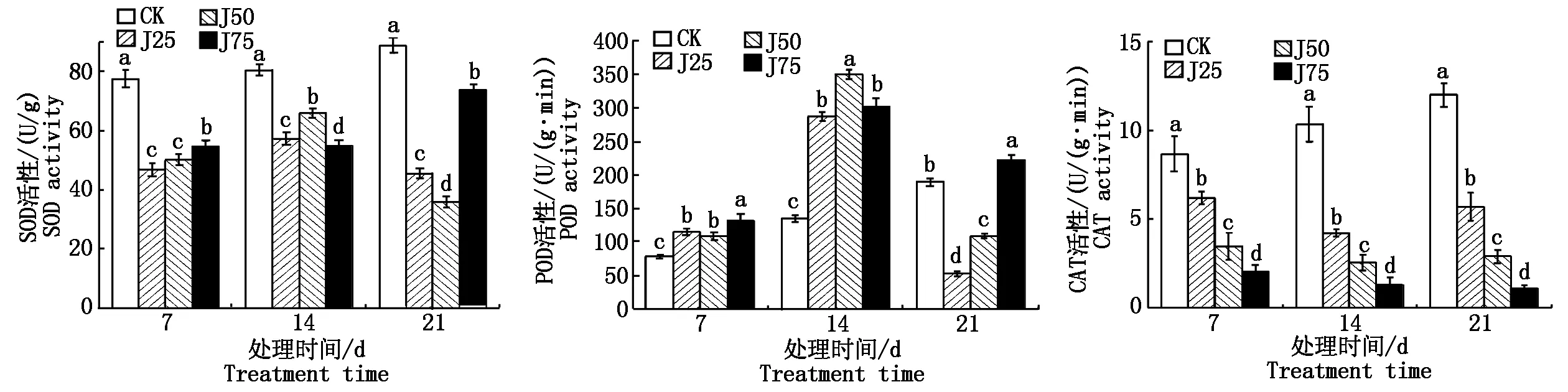
图6 碱胁迫对甘蓝型油菜叶片抗氧化酶活性的影响Fig.6 Effect of alkali stress on antioxidant enzyme activity in leaves of Brassica napus L.
如图7所示,在处理第7天,处理组SOD活性要高于对照组,随着复合盐碱溶液浓度及处理天数增加,SOD活性降低;复合盐碱处理提高了POD活性,与对照相比,高盐碱(YJ75)处理21 d后的POD含量提高了2.26倍,差异显著(P<0.05);处理组中,CAT活性随着复合盐碱的升高而呈先升后降的趋势,而随着胁迫时间的持续,CAT的活性呈先降后升的趋势,处理组中CAT活性均低于对照组,差异显著。

图7 盐碱胁迫对甘蓝型油菜叶片抗氧化酶活性的影响Fig.7 Effect of saline-alkali stress on antioxidant enzyme activity in leaves of Brassica napus L.
各项生理指标结果均表明,盐碱胁迫对油菜幼苗的伤害程度大小表现复合盐碱>碱>盐。
2.3 甜菜碱合成途径中关键酶基因表达量
由图8可知,复合盐处理7 d,高盐(Y75)处理后的CMO基因表达量达到最高值,为空白对照的3.14倍,而在处理14,21 d,CMO基因的表达量随着盐浓度的升高而上升,差异显著(P<0.05);图9,10可以看出,复合碱与复合盐碱溶液显著提高了CMO基因的表达量;在复合盐、碱、盐碱溶液处理21 d后,高盐(Y75)、高碱(J75)、高盐碱(YJ75)处理组中CMO基因相对表达量分别提高7.21,9.23,13.76倍,差异显著(P<0.05);同浓度的复合盐碱处理后的叶片中的CMO基因表达量要高于复合盐与复合碱处理组。
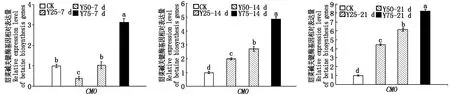
图8 盐胁迫对合成甜菜碱关键酶基因相对表达量的影响Fig.8 Effect of salt stress on relative expression of key enzyme genes in betaine synthesis

图9 碱胁迫对合成甜菜碱关键酶基因相对表达量的影响Fig.9 Effect of alkali stress on relative expression of key enzyme genes in betaine synthesis

图10 盐碱胁迫对合成甜菜碱关键酶基因相对表达量的影响Fig.10 Effect of saline-alkali stress on relative expression of key enzyme genes in betaine synthesis
在油菜幼苗受到盐碱胁迫后,关键酶基因CMO的相对表达量随着复合盐、碱、盐碱溶液浓度的升高而上升,且胁迫时间越长差异越显著。
2.4 甜菜碱积累量
如图11所示,甜菜碱的含量随复合盐、碱、盐碱的浓度及胁迫天数的增加而上升,且差异显著(P<0.05)。在高浓度的盐(Y75)、高浓度的碱(J75)、高浓度的盐碱(YJ75)胁迫最高天数(21 d)时,油菜叶片中的甜菜碱的积累量达到了最高值,分别为7.99,9.61,15.00 mg/g。
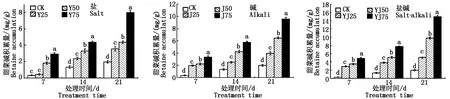
图11 甘蓝型油菜叶片中甜菜碱含量Fig.11 The betaine content in leaves of Brassica napus L.
3 结论与讨论
土地盐碱化是世界范围内普遍存在的一个问题,严重影响着植物的生长和发育[22]。种子萌发代表着植物生命的开始,在种子萌发阶段,其发芽率与植物耐盐碱能力密切相关。本试验中,油菜种子的发芽率随着各溶液浓度的升高呈下降趋势,其中中高浓度的复合盐碱胁迫(YJ50、YJ75)对油菜种子的伤害最大,而复合盐溶液对种子发芽率的影响最小,说明盐碱胁迫对种子的伤害程度远大于单盐、单碱对种子的伤害。
可溶性糖和脯氨酸可维持细胞代谢平衡、细胞内稳态和细胞膜稳定性,当植物受到盐碱胁迫时,脯氨酸、可溶性糖、甜菜碱等渗透调节物质发挥了极其重要的作用[23]。本试验中,脯氨酸、可溶性糖、甜菜碱均随着盐碱溶液浓度及胁迫天数的增加而呈上升的趋势。
本试验中,叶绿素的含量对中低浓度复合盐溶液(Y25、Y50)表现为随处理时间延长呈现先升高后降低的趋势,可能是因为此时油菜叶片中的脯氨酸、可溶性糖、甜菜碱等渗透调节物质发挥了重要作用,修复了油菜叶片所受到的损伤,有效抑制了叶绿素的降解,在经过复合碱、复合盐碱及高复合盐(Y75)溶液处理后,叶绿素含量随着处理浓度与处理时间的升高而下降。
有研究表明,丙二醛含量是随着盐浓度或胁迫时间的增加呈上升趋势[24],这一观点与本研究结果一致。本试验中,盐碱胁迫显著提高了油菜叶片丙二醛含量,而在盐碱胁迫7 d,丙二醛含量变化差异较小,说明短时间的盐碱胁迫对甘蓝型油菜幼苗所产生的伤害较低,可能是因为当油菜幼苗因丙二醛的积累而受到活性氧的伤害时,SOD、CAT、POD作为酶类保护体系开始增加,其中POD发挥作用最大。
甜菜碱合成途径中的关键酶基因CMO的相对表达量随着盐碱溶液的浓度升高以及胁迫时间延长而上升,CMO会将胆碱氧化成甜菜碱,因此甜菜碱的含量也会随之升高,在复合盐、碱、盐碱溶液处理21 d后,与对照组相比,高盐(Y75)、高碱(J75)、高盐碱(YJ75)处理组中CMO基因的相对表达量显著提高,且此时甜菜碱的积累量也达到了最高值,因此在甘蓝型油菜受到盐碱胁迫时,甜菜碱会大量积累以减轻油菜所受的伤害。
综上所述,复合盐、碱、盐碱溶液对甘蓝型油菜的伤害程度大小表现为复合盐碱>碱>盐;低浓度复合盐溶液对甘蓝型油菜种子的发芽率影响较低,且短时间复合盐处理对叶绿素含量有所增加,说明适当浓度盐溶液对甘蓝型油菜的生长具有一定的促进作用;盐碱溶液会提高甘蓝型油菜幼苗中的可溶性糖、脯氨酸、甜菜碱等渗透调节物质以降低盐碱胁迫对油菜所带来的伤害;丙二醛的积累会使甘蓝型油菜细胞器发生损伤,影响其生长,此时抗氧化酶起到了重要作用:清除活性氧,提高植物的耐盐碱性,其中以POD发挥的作用最大;CMO作为甜菜碱合成途径中的关键酶基因,它的增加促进了甜菜碱的积累,而抵御甘蓝型油菜所遭受的盐碱胁迫所带来的伤害。以上结果为甘蓝型油菜耐盐碱品种选育和改良提供了一定的理论基础。
猜你喜欢
金桥(2021年11期)2021-11-20 06:37:14
现代畜牧科技(2021年4期)2021-07-21 06:12:54
西藏农业科技(2019年3期)2019-11-04 00:35:14
西藏农业科技(2019年1期)2019-07-25 00:37:02
西藏农业科技(2018年4期)2018-04-25 06:39:28
西藏科技(2016年8期)2016-09-26 09:00:55
西南农业学报(2016年6期)2016-04-16 05:12:46
中国麻业科学(2015年5期)2015-12-28 06:22:10
中国洗涤用品工业(2015年8期)2015-02-28 19:02:48
无机化学学报(2014年9期)2014-02-28 17:33:08
