
中华绒螯蟹养殖群体与野生群体的种群遗传学研究
2022-07-09 07:25周华兴段国庆胡玉婷
农学学报 2022年6期
周华兴,段国庆,江 河,凌 俊,胡玉婷
(安徽省农业科学院水产研究所,水产增养殖安徽省重点实验室,合肥 230031)
0 引言
中华绒螯蟹(Eriocheir sinensis),俗称河蟹,隶属节肢动物门、甲壳纲、十足目、方蟹科、绒螯蟹属,是重要水产养殖品种,其营养价值丰富,口味鲜美,深受人们喜爱。20世纪70年代末,中华绒螯蟹内陆水域人工育苗技术难关攻克,增养殖业有了长足的发展[1]。尤其是在20世纪90年代,养殖产业突飞猛进,极大的推进了中华绒螯蟹资源的开发与利用。
中华绒螯蟹是典型的降海洄游型生物,其幼体在江、河等淡水中生长发育至成熟个体后,顺水而下至入海口等咸水中繁殖[2]。长江入海口浅滩广阔、资源丰富,是中华绒螯蟹天然的繁育场[3]。野生中华绒螯蟹在长江口繁殖后,逆流而上,在长江流域生长发育,形成特有的长江品系,沿江流域也随之开展了大量的河蟹养殖。然而,养殖过程中大规模跨地域的引种以及养殖群体的逃逸都可能造成中华绒螯蟹野生群体的种质混杂[4-5]。王海华[6]采用线粒体控制区序列对长江水系7 个野生群体的遗传多样性分析结果表明,长江水系中华绒螯蟹各个群体间分化不显著,没有出现与地理分布相对应的谱系结构。李晶晶等[7]利用10个微卫星分子标记分析了中华绒螯蟹3个不同水系人工选育群体和1个野生群体的遗传分化水平,结果表明4个群体的遗传分化程度处于中低水平。学者认为遗传漂变的缺失和显著的同质化是造成种质混杂的内在因素[8]。
为探究中华绒螯蟹养殖群体对野生群体的遗传多样性影响,本研究采集长江流域安徽段野生群体样本以及邻近的养殖群体样本,结合相关分子学手段,探究长江中华绒螯蟹种质资源状况以及资源混杂程度,以期为中华绒螯蟹资源的科学保护、合理利用以及相关产业政策的制定提供理论依据。
1 材料与方法
1.1 样本采集
研究于2016年采集了江流域安徽段中华绒螯蟹4个主要养殖区,分别是芜湖县、无为县、当涂县、宣州区(如图1、表1 所示)的养殖样本共计140 尾;采集长江流域芜湖至马鞍山段中华绒螯蟹野生样本30 尾。样本采集后,取其腿部肌肉保存于-20℃下。实验于2016年12月—2018年3月,在水产增养殖安徽省重点实验室进行。
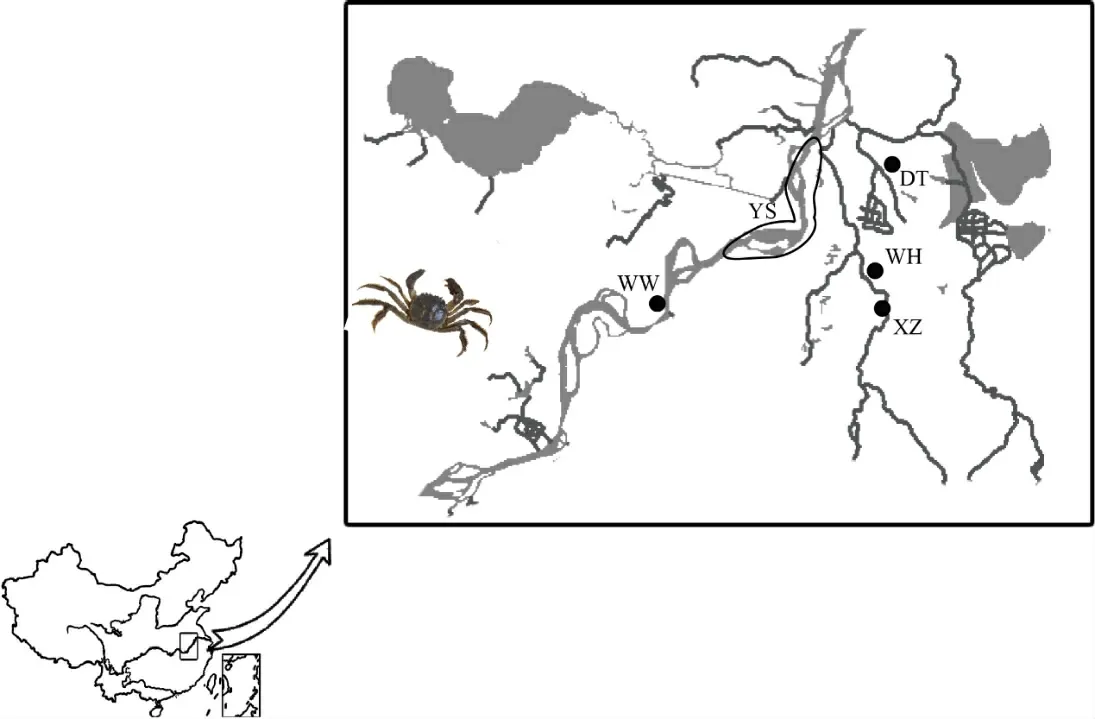
图1 样本采集点分布图
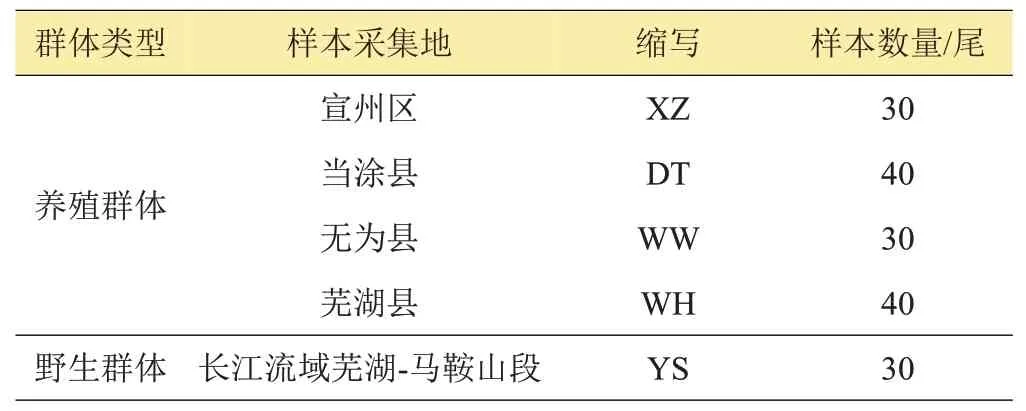
表1 样本采集信息表
1.2 基因组DNA提取与PCR扩增
取中华绒螯蟹肌肉组织,采用Sambrook法提取基因组DNA[9]。参照NCBI 上已有的中华绒螯蟹线粒体DNA(mtDNA)全序列数据(登录号:KP126617),使用Oligo7[10]设计线粒体COII基因的引物(表2),并委托上海生工生物有限公司合成。
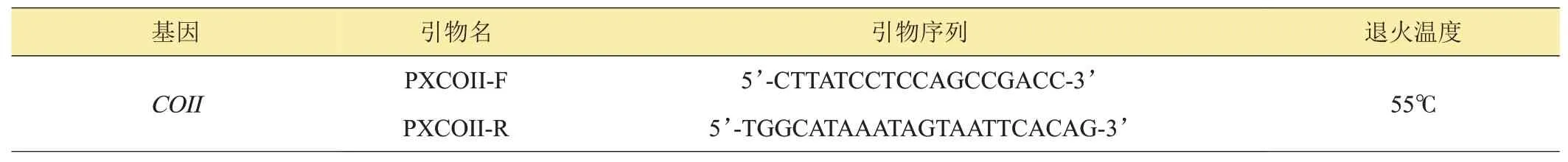
表2 中华绒螯蟹线粒体基因COII引物
PCR 反应体系:包含200 ng 的模板DNA,10×Buffer 缓冲液(TaKaRa)5 µL,dNTP(2.5 mM)3 µL,MgCl2(2.5 mol/L)4µL,上下游引物(5µmol/L)各2µL,TaqDNA聚合酶0.75 U,补充无菌超纯水至50µL。
PCR 反应条件:96℃预变性1 min;94℃变性45 s,55℃退火45 s,72℃延伸1 min,共34个循环;最后72℃延伸10 min。PCR 产物经1%的琼脂糖凝胶电泳检测后,预期长度的线粒体DNA 片段通过DNA 纯化试剂盒(DNA Gel Extract Purification Kit,TaKaRa)回收后,委托上海生工生物有限公司双向测序。
1.3 序列分析
测序所得序列通过Seqman 软件辅助人工进行拼接组装。序列通过MAFFT v.7[11]在线比对后,使用DNAsp v.6.11[12]计算多态位点、单倍型数目、单倍型多样性(Haplotype diversity,Hd) 和核苷酸多样性(Nucleotide diversity,Pi)。使用Arlequin v.3.11 进行分子变异分析(Analysis of Molecular Variance,AMOVA)[13]。使用 Mega v.7.0[14]基于邻接法(Neighbour joining,NJ)构建单倍型分子系统树。采用Network 5000 基于中介网络法(Median Joining,MJ)绘制单倍型网络关系图[15]。
2 结果与分析
2.1 序列特征
测序获得170条长度为799 bp的中华绒螯蟹线粒体COII基因序列,共检测到突变位点38个,其中简约信息位点10个,单突变位点28个。基因片段中,T、C、A、G四个碱基的平均含量分别为35%、19.4%、31.2%、14.4%。A+T 的含量为66.2%,大于G+C 的含量33.8%。基因序列出现了明显的反G偏倚(Anti-G bias)现象[16]。
2.2 单倍型进化分析
中华绒螯5个群体中共检测出12种单倍型,其中XZ 群体有7 种单倍型,WH 群体有6 种单倍型,DT 和WW群体均有5种单倍型,YS群体有4种单倍型(见表3)。12种单倍型中,Hap 6、11、12为群体特有单倍型,其余9个单倍型均为群体共享单倍型。

表3 中华绒螯蟹5个群体单倍型和遗传多样性信息
选择日本绒螯蟹COII序列(登录号:NC_011597)作为外类群,构建单倍型系统发育树,结果表明,Hap12独立成支,其余11个单倍型聚为一支。单倍型网络关系分析结果表明,Hap 1 和2 包含了所有的群体,且Hap 2的共享个体数最多,为98个。除Hap 12外(突变步数为30),其余11个单倍型间的突变步数均为1(图2)。
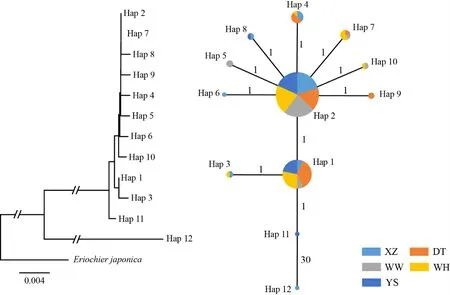
图2 12个单倍型系统发生树(左)和中介网络关系图(右)
2.3 遗传多样性和分子变异分析
种群遗传多样性分析结果表明DT群体单倍型多样性最高,XZ 群体核苷酸多样性最高;WW群体的单倍型多样性和核苷酸多样性均最低,分别为0.4090和0.0006。野生中华绒螯蟹YS 群体的单倍型多样性为0.5450,核苷酸多样性为0.0008,仅高于WW群体。
分子变异分析结果表明,群体内的分子变异为96.54%,远高于群体间的变异(3.46%,P<0.05)(表4)。群体间的平均遗传分化指数Fst值为0.0346,说明多个群体间没有出现遗传分化[17]。
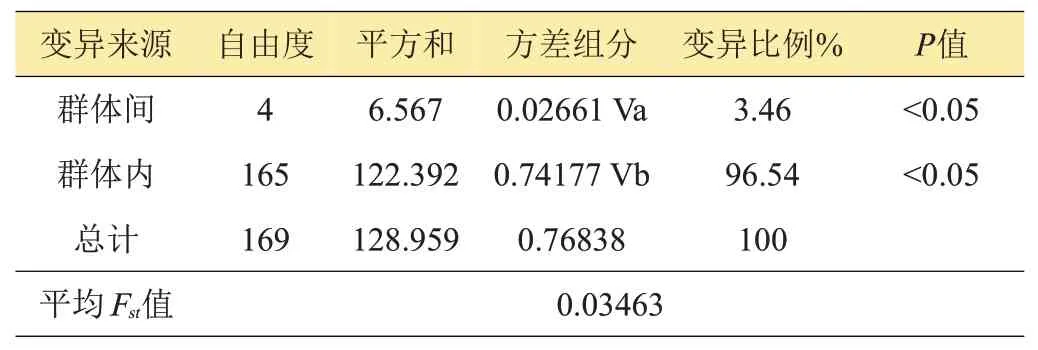
表4 种群分子变异分析
3 讨论
3.1 长江野生中华绒螯蟹遗传多样性
遗传多样性是生物群体抗逆和进化的潜在动力,也是种群适应外界环境能力最直观的反映[18]。群体的遗传多样性水平往往受到多种因素的影响,如谱系分选、特殊的生活史等[19-20]。中华绒螯蟹作为重要的水产品,具有极高的经济价值,其自然群体受到人类活动的影响较大。
中华绒螯蟹线粒体COII基因序列中,A+T的含量为66.2%,G+C 的含量为33.8%,表现出明显的碱基组成偏倚。有学者基于线粒体COI基因序列的中华绒螯蟹野生群体遗传多样性分析[21]中,A+T 的含量为61.7%,G+C 的含量为38.3%,这与本研究的结果相印证。本研究中,长江芜湖-马鞍山江段的野生中华绒螯蟹遗传多样性水平较低(Hd=0.5450;Pi=0.0008),低于2008 年报道的长江南京江段中华绒螯蟹群体遗传多样性(Hd=0.8433;Pi=0.0048)[22]和2016 年报道的长江口中华绒螯蟹群体遗传多样性(Hd=0.8333;Pi=0.0077)[21]。这说明长江流域中华绒螯蟹群体近年来出现了种质退化现象。王海华[6]通过系统分析长江中华绒螯蟹资源变动以及资源危机爆发原因,提出长江中华绒螯蟹主要致危因素有水利工程、湖泊围垦、水质污染和酷鱼滥捕。野生中华绒螯蟹群体遗传多样性的下降弱化了其对环境的适应能力,最终可能导致野生资源的衰退。
3.2 中华绒螯蟹养殖和野生群体种群遗传分化
采样调研结果表明,4 个养殖群体的中华绒螯蟹繁殖亲本来源差异较大,其中DT 群体的亲本主要来自当涂县石臼湖;WW群体的亲本主要来自常州市长荡湖;WH 群体的亲本主要来自如东县;XZ 群体的养殖品种为选育新品种‘长江2 号’,其亲本来自莱茵河水系。然而,多个群体间并未检测到明显的遗传分化。研究中,单倍型网络进化分析和系统发育分析结果表明,Hap 1-11 形成一支,为并系关系;Hap 12 形成单独的进化支,可能是莱茵河水系中华绒螯蟹的遗传特征;5 个群体未出现明显的谱系分化。群体间的平均遗传分化指数Fst值为0.0346,说明多个群体间没有出现遗传分化。AMOVA 分析结果表明,不同群体间的遗传变异远小于群体内个体间的遗传变异。
5 个群体间没有明显的遗传差异,但养殖群体亲本来源差异大,说明养殖群体和野生群体间存在严重的种质混杂。Chang 等[4]也曾报道过长江野生中华绒螯蟹种质可能受其他水系污染。研究中采集的养殖场毗邻长江干流,在雨水充足的季节,养殖场易被洪水淹没。而相比其他水产动物,中华绒螯蟹的运动能力强[23],可借助水流从养殖场四散爬出,最终汇入长江干流。这种养殖逃逸可能是造成长江中华绒螯蟹种质混杂的主要原因。
4 结论
本研究采集长江流域安徽段中华绒螯蟹野生群体样本以及邻近的养殖群体样本,基于线粒体COII基因序列,开展了种群遗传学分析。结果表明,长江流域中华绒螯蟹养殖群体与野生群体间未出现显著的遗传分化,野生群体基因库可能受到养殖群体的影响,种质混杂现象严重。与前人研究结果相比,近年来中华绒螯蟹长江野生群体遗传多样性下降,资源衰退,建议加大野生资源的保护力度,坚持保护优先原则。
猜你喜欢
世界中医药(2022年18期)2022-11-25
南京医科大学学报(自然科学版)(2022年8期)2022-11-22
重庆理工大学学报(自然科学)(2022年9期)2022-10-26
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国种业(2022年1期)2022-01-27
中国种业(2022年1期)2022-01-27
现代农业科技(2009年19期)2009-03-20
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26
