
植物多糖的益生作用及其影响因素研究进展
2022-07-02 03:49杨斯惠曹亚楠任远航彭镰心时小东
食品科学 2022年11期
杨斯惠,向 月,曹亚楠,*,任远航,彭镰心,*,时小东
(1.成都大学 农业农村部杂粮加工重点实验室,四川 成都 610106;2.四川省杂粮产业化工程技术研究中心,四川 成都 610106)
多糖是由超过10个单糖基经过-或-糖苷键聚合、脱水形成的天然高分子化合物,主要分布于动物、植物、藻类及微生物中,可以根据其化学成分、在植物体内的功能、来源、溶解性等进行分类。根据来源的不同,可将多糖划分为动物多糖、植物多糖和微生物多糖(真菌多糖、细菌多糖)。植物多糖作为植物体的重要组成物质,与蛋白质、核酸、脂类并列为植物生命体四大基本物质。常见的植物多糖有淀粉、纤维素、葡聚糖、果胶、果聚糖等。随着越来越多的科研工作者投入对植物多糖的研究中,其在加工、结构和生物活性方面受到关注。研究表明,植物多糖在抗氧化、抗肿瘤、抗肝损伤、降血糖、益生作用等方面具有显著优势。其中,益生作用是多糖生物活性的一个重要方面。
根据推测,人体内存在约500~1 000种细菌,这些微生物总共构成了100万亿个细胞,是自身细胞数量的10 倍,而大多数微生物在肠道生存。这些在人和动物肠胃中生活的微生物统称为肠道菌群。健康个体的肠道菌群中,大多数微生物分布在厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、螺旋体门(Spirochaetes)、梭杆菌门(Fusobacteria)、蓝藻菌门(Cyanophyta)、疣微菌门(Verrucomicrobia)以及黏胶球形菌门(Lentisphaerae)这9 类菌门中,其中至少90%的细菌属于厚壁菌门和拟杆菌门。绝大多数微生物群的定植始于生命早期,在新生儿出生时和出生后不久,新生儿接触了母体和环境中的微生物,同时肠道菌群的建立启动。婴儿发育、肠道形态和饮食变化等因素影响婴儿肠道菌群的变化,结果表现为兼性厌氧菌数量减少,严格厌氧细菌逐渐增加成为优势菌种。哺乳期结束后,肠道菌群趋于稳定,健康人体内形成终生的菌群特征。正常机体内,肠道菌群能够代谢宿主自身不能降解的多糖,增强肠道上皮屏障功能,减轻炎症反应,调节免疫反应,抑制致病菌群生长的功能。然而当微生物受到环境、年龄、饮食、生活方式、抗生素等各类因素影响时,菌群动态平衡被打破,进而导致菌群混乱。而通过在饮食中补充多糖调节肠道微生物区系也逐步被人们认识到是有效方法。
据报道,植物多糖(淀粉除外)不能被人体直接消化,但可作为益生元通过抑制病原体和刺激免疫系统来维持机体胃肠道健康,被肠道菌群分解利用,生成具有生物活性的产物(如短链脂肪酸(short-chain fatty acids,SCFAs)),调节肠道微生物组成和活性,进而促进人体健康。而多糖的生理功能又与其单糖组成、分子质量、分子形状和链构象等结构密切相关。当前随着对植物多糖生物活性深入研究,其在医学、食品、生命科学等领域广泛运用,成为国内外研究热点。本文系统论述植物多糖与其益生作用的关系以及影响植物多糖结构的因素,旨在为植物多糖的精准化开发利用提供参考。
1 植物多糖的益生作用
植物中非淀粉类多糖在唾液、胃液和肠液等消化液中不能被消化或完全消化,其中不能被宿主自身酶类催化水解的植物多糖进入肠道,成为人体内肠道微生物区系的主要碳源。利用碳水化合物活性酶类(carbohydrate-active enzymes,CAZymes)的交互降解作用是肠道菌群利用植物多糖的主要途径。肠道菌群编码的CAZymes主要包括糖苷水解酶(glycoside hydrolases,GHs)、糖基转移酶(glycosyltransferases,GTs)、多糖裂解酶(polysaccharide lyases,PLs)、碳水化合物酯酶(carbohydrate esterases,CEs)和碳水化合物结合模块(carbohydrate binding modules,CBM)。据报道,肠道菌群分解利用多糖的能力与其编码的CAZymes数量密切相关,同一类型但源于不同菌种的CAZymes活力存在差异,因此,不同菌属或菌株分解植物多糖的能力可能不同,不同的植物多糖来源还会影响降解微生物中基因表达的特异性。在结肠环境中,植物多糖在肠道菌群的作用下通过不同的降解系统和转运系统被降解成单糖或低聚糖,随后这些单糖或低聚糖被转运至胞内进一步降解和发酵,产生气体(氢气、二氧化碳和甲烷)、热量和SCFAs。SCFAs是肠道微生物群酵解植物多糖的产物中研究最多的。醋酸、丙酸和丁酸是粪便中最常检测到的SCFAs,是不同细菌的代谢产物。许多细菌可以产生醋酸,但丙酸和丁酸通常是由特定的细菌产生的。在胃肠道环境中,丁酸的主要产生菌是厚壁菌门(如毛螺菌科和普氏杆菌),而丙酸是由类杆菌、阴性杆菌和一些梭状芽孢杆菌产生的。SCFAs在人体内如何通过碳水化合物代谢途径获得以及发挥何种生理功能已有较为详细的叙述。研究表明,植物多糖对肠道菌群组成具有调节作用,能够促进有益细菌生长,产生SCFAs,降低粪便pH值,抑制有害菌生长(图1)。
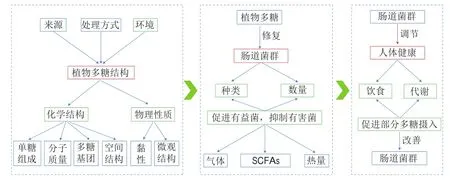
图1 植物多糖对肠道菌群和人体健康的调节作用Fig.1 Regulatory effect of plant polysaccharides on intestinal flora and human health
由表1可知,竹笋多糖、枇杷叶多糖、黄芪多糖、党参多糖、白术多糖、大粒车前子多糖、菊粉型果聚糖和枸杞多糖均能促进双歧杆菌的增殖,果胶多糖和枇杷叶多糖可以显著增加巨球藻属和脱硫弧菌属的数量,燕麦多糖和龙眼多糖则能促进干酪乳杆菌的增殖。这些益生菌相对丰度改变影响着代谢产物SCFAs的生成。植物多糖的来源具有多元性,而不同的植物多糖针对不同的益生菌作用不同,这可能与多糖高度复杂的结构、物理特性密切相关。

表1 植物多糖的益生作用Table 1 Prebiotic effects of plant polysaccharides

续表1
2 植物多糖结构与肠道菌群的关系
2.1 化学结构
植物多糖是结构形式复杂的高分子化合物。植物多糖可以分为一级结构、二级结构、三级结构和四级结构。一级结构包括单糖的组成、糖苷键的连接方式和顺序、异头碳的构型等;二级结构是指多糖主链之间通过氢键等次级键而形成的聚合体结构;类似于其他生物大分子,三级和四级结构是指在多糖的二级糖单位结构的基础上,通过非共价作用在有序空间里产生的规则构象。近年来,从植物中提取得到的多糖在医学、食品、生命科学等领域引起了广泛关注,其具有多种生理活性如抗肿瘤、免疫调节、降血糖、益生作用等。这些生理活性受其分子质量、链构象、糖苷键类型等结构的影响很大。
2.1.1 单糖组成
植物多糖的单糖组成与益生元活性具有相关性。单糖组成能够影响多糖的链结构与高级结构,而高级结构是影响多糖生物活性强弱的一个重要因素。由表1可知,大部分具有益生作用的多糖都具有半乳糖、阿拉伯糖、葡萄糖、鼠李糖、半乳糖醛酸和甘露糖,还有少数具有木糖和果糖。王轶帆等报道了由不同单糖残基组成的杂多糖——龙眼多糖与由相同单糖残基组成的同多糖——燕麦多糖的结构特征及其益生活性比较,发现单糖组成更为复杂的龙眼多糖相较于主要由葡萄糖和少量甘露糖组成的燕麦多糖促益生菌增殖效果更佳,推测可能是由于杂多糖作为碳源被益生菌消耗较慢,有利于维持益生菌活菌数稳定。以半乳糖和半乳糖醛酸为基本构成单元的低聚半乳糖具有多种生物活性,同时具有较强的耐酸性和耐热性,不会在高温杀菌的加工过程中被破坏,也很少因人体胃酸分解而失去活性。其中,低聚半乳糖的益生活性与地锦草表现出的治疗腹泻等功效相符,李胜男推测由大量半乳糖组成的地锦草多糖可能也具有低聚半乳糖的益生活性。Zhu Rugang等报道了猕猴桃果实中果胶低聚糖(pectin oligosaccharides,POSs)与抗糖化潜力的构效关系,发现POSs的益生元活性与分支度((阿拉伯糖)∶(鼠李糖)和(半乳糖)∶(鼠李糖))以及中性糖含量呈正相关,特别是半乳糖和阿拉伯糖。从阿月浑子壳和竹笋中提取的由阿拉伯糖、半乳糖和木糖组成的多糖是潜在益生元也印证了多糖的单糖组成与益生元活性具有相关性这一观点。
2.1.2 分子质量
分子质量在一定范围内的多糖具有生物活性,分子质量过大或过低的多糖一般不具有明显的生物活性。究其原因,是因为活性多糖通过跨越细胞膜进入生物体内发挥生物学活性,而分子质量的大小影响了多糖的溶解性和黏度,与多糖跨越细胞膜障碍相关,分子质量太大不利于实现这个过程。与分子质量278、354 kDa的阿拉伯木聚糖相比,分子质量66 kDa的阿拉伯木聚糖对于益生菌生长具有更好的增殖作用。Chen Guangjing等报道了不同提取方法对竹笋多糖益生元活性影响的研究,探究提取工艺对竹笋多糖的体外消化率、益生元活性和理化性质的影响。结果发现,超声辅助提法得到的竹笋多糖(U-CPS)和酶解法得到的竹笋多糖(E-CPS)的分子质量均显著低于另外3种多糖。同时体外研究发现,分子质量和粒径较小的U-CPS和E-CPS可能更利于被细菌细胞转运和代谢,刺激SCFAs的生成,从而产生更好的益生作用。Huang Fei等发现分别采用热水、超细粉碎和超细粉碎辅助酶法提取得到的龙眼果肉多糖LP-H、LP-S和LP-SE对益生菌增殖有积极作用,且分子质量较小的LP-SE对益生菌增殖作用最强。Fu Yuan等利用7种方法(热水提取、加压水提取、高速剪切均质提取、微波辅助提取、超声辅助提取、超声波辅助酶法提取和超声-微波辅助)提取枇杷叶多糖,其中超声辅助提取和超声-微波辅助提取的枇杷叶多糖具有较强的益生作用,这可能与其具有较低的分子质量和黏度有关。张宇等通过灌胃盐酸林可霉素建立肠道微生态失调小鼠模型,然后利用不同分子质量的黄芪多糖进行治疗,结果表明黄芪多糖分子质量在6~10 kDa范围内有利于“扶持”有益菌,抑制有害菌,调节小鼠肠道微生态菌群平衡效果最佳。
2.1.3 多糖基团
多糖基团数量、位置和种类的改变会导致生物活性的改变。相关研究表示,黄秋葵花多糖能够显著促进大肠杆菌发酵液中乙酸和己酸的产生。而进行羧甲基化、硫酸酯化和乙酰化结构修饰后,与黄秋葵花多糖相比,修饰多糖显著抑制大肠杆菌发酵液中乙酸、丁酸和己酸的产生,这可能是因为修饰多糖不能被大肠杆菌酵解发酵产生SCFAs。王亚莉以葡聚糖硫酸钠诱导的小鼠溃疡性结肠炎为模型,研究松花粉多糖和硫酸酯化松花粉多糖对葡聚糖硫酸钠组小鼠肠道菌群的影响,发现两种多糖在调节肠道菌群平衡方面都起到积极作用。王鑫以菜籽为原料,经提取、分离和纯化得到均一多糖R/MIN1-a,然后对R/MIN1-a分别进行硫酸化修饰、乙酰化修饰和羧甲基化修饰,研究R/MIN1-a和修饰后多糖对肠道益生菌体外增殖活性的影响。结果表明,与R/MIN1-a相比,硫酸化修饰产物对嗜酸乳杆菌、婴儿双歧杆菌、青春双歧杆菌和两歧双歧杆菌的增殖作用更为明显。Nuerxiati等对广布红门兰多糖进行硫酸化、乙酰化、磷酸化和羧甲基化修饰,发现不同的改性多糖对不同的益生菌增殖效果不同;在保加利亚乳杆菌增殖效果的比较中,磷酸化多糖表现最佳,乙酰化多糖对青春双歧杆菌的生长促进效果最好,硫酸化多糖则保持适度稳定的益生菌能力。
2.1.4 空间结构
特定的空间结构是发挥多糖生物活性的必要条件。作为影响多糖生物活性强弱的一个重要因素,多糖的空间结构复杂,是糖链之间或者糖链内部氢键、范德华力、色散力和疏水性等非共价键共同作用的结果,并具有多种构象,例如单螺旋、双螺旋、三螺旋和无规卷曲等。在多糖空间结构的分析测定上,常用的方法有刚果红实验、X射线衍射和原子力显微镜分析等。一般认为,多糖具有规则的空间结构,则生物活性也强。研究发现,多糖生物活性的基础是-(1→3)--葡聚糖的单股螺旋构象。例如,-(1→3)--葡聚糖发挥免疫增强活性主要依赖于单螺旋构象和位于螺旋外表面的亲水性基团的存在,三股螺旋结构对其活性的发挥来说并不是必须的。而Maeda等认为,三股螺旋构型对有些食用菌-(1→3)-葡聚糖的免疫刺激和抗肿瘤活性有重要作用。虽有较多文献报道多糖空间结构与活性的关系,也取得了重要进展,但由于植物多糖的复杂性和高度多态性,许多精细结构仍未得到准确解析,相关研究结果仍存争议。植物多糖益生作用虽已被证实,但其空间结构与益生活性的关系鲜有报道,制约了植物多糖的精准化开发利用。
2.2 物理性质
2.2.1 黏度
多糖的黏度由溶解度和分子质量所决定。而黏度与非淀粉多糖中的单糖组成和键型没有相关性。在较低浓度下,多糖会与水分子直接结合,黏度随之增加;在较高浓度下,多糖相互结合和共价或非共价键连接形成网状物,更易于增加肠道内容物的黏度。一般来说,分子质量越大、分子结构越复杂的多糖增加消化道中液体黏度的作用愈明显。而研究表明,水溶性高和黏度低的多糖可以更容易、快速和完全地被益生菌利用。
2.2.2 微观结构
多糖的微观结构受单糖组成的影响,随单糖组成复杂程度、数量和连接方式的改变,因此也会有所不同。用扫描电子显微镜观察多糖的形态可以有效地分析多糖的结构和形态,包括大小、分子形状和孔隙率。Xu Shuying等分析了人类肠道菌群对海坛紫菜多糖的微生物分解代谢,通过扫描电子显微镜观察发现,海坛紫菜多糖的结构可能由较不易被肠道微生物降解利用的外层和容易被肠道细菌降解的内层组成,从而增加了SCFAs的水平。
3 影响植物多糖结构的因素
植物多糖益生作用受到学者们广泛关注,以植物多糖为原料的益生产品开发成为热点。尽管其益生机制尚未明晰,植物多糖结构与物理性质关系到其活性强弱。多糖的结构受到来源、处理方式、环境、温度和其他因素的影响(图1)。认识改变植物多糖结构的关键因素并合理利用,可为植物多糖开发利用提供重要参考。
3.1 提取方法
针对植物多糖的提取,常用的方法有热水浸提、酸提法、碱提法、醇提法、超声波提法、酶解法等。
热水浸提是最为经济、便捷和常见的多糖提取方法,能够较好保存分子结构,但提取耗时长,效率低,得到的多糖大多是中性多糖,还有可能降解多糖。酸提法原理是在一定温度、时间和pH值条件下,细胞膨胀并破坏细胞壁,植物多糖从中释放。酸提法具有提取率高、纯度高、耗时短的优点;缺点是具有特异性,不适用于一般植物多糖的提取,并破坏糖苷键。碱提法原理与酸提法相似,适合提取含有糖醛酸的多糖及酸性多糖,有利于提取率的提高。酶解法是通过酶对细胞壁通透性的改变,减少溶剂提取时来自细胞壁、细胞膜和细胞间质的阻力,使其软化、膨胀、破裂,从而使内容物流出。此法具有反应条件温和、工艺简单、生物活性高等优点,但与传统水提醇沉法相比,成本较高。
张红运研究了不同提取工艺对大豆种皮多糖结构的影响,利用微波辅助提取法和热水浸提法提取多糖,发现微波辅助提取有利于大分子多糖的提取,但具有多糖分子质量分布范围较广、均一性差等缺点,同时还会降低分支链分子质量。马智玲分别采用酸法、酶法和盐法对马铃薯果胶进行提取,结果表明,不同提取方法下,果胶的单糖组成和分子质量不同,提取剂的性质和沉析方式也会对果胶的结构造成影响。Chen Guangjing等探究了提取工艺对竹笋多糖理化性质的影响。结果发现,不同提取工艺下,5种多糖的单糖组成相同,但各单糖的含量不同;超声辅助提法得到的竹笋多糖和酶解法得到的竹笋多糖的分子质量均显著低于另外3种多糖。Huang Fei等采用热水、超细粉碎和超细粉碎辅助酶法提取龙眼果肉多糖,结果表明,3种提取方法对多糖的化学组成和结构均有一定影响。与热水、超细粉碎法相比,超细粉碎辅助酶法提取多糖的表观黏度、粒径、分子质量最低。
3.2 分离、纯化
多糖经过提取后一般还夹杂着蛋白质、色素、小分子杂质等物质。对于多糖结构和生物活性的研究,蛋白质的存在会造成干扰。因此,选用对多糖结构和生物活性不造成破坏的脱除方法很重要。常用Sevag法、三氟三氯乙烷法、阴离子交换树脂法、酶解法等除去蛋白质。Sevag法是脱除蛋白质的经典方法,原理为蛋白质在氯仿等有机溶剂中发生变性。Sevag法的特点为条件温和、效率低、操作琐碎和得率不高等。与Sevag法相比,三氟三氯乙烷法脱除蛋白的效率较高,但溶剂易挥发,不宜大规模生产应用。阴离子交换树脂法可调节溶液pH值使蛋白质形成阴离子,采用阴离子交换树脂作为流动相,具有去除效果好、成本低、条件温和等特点。常用活性炭吸附法、乙醇洗涤法、过氧化氢法等除色素。除小分子杂质则主要是用透析法和超滤法。分离除杂后,利用各种多糖的溶解度不同及电荷密度差异进行纯化得到单一的多糖物质。常用的纯化方法有分级纯化法、凝胶柱层析、超滤法等。任益平以藜麦为原料采用超声波辅助提取、AB-8大孔树脂吸附、DEAE纤维素分离和Sephadex G-100凝胶柱纯化后得到4个组分多糖,再通过傅里叶变换红外光谱分析、核磁共振波谱分析、刚果红实验和微观结构分析研究分离纯化对多糖结构的影响。结果表明,在分离纯化的过程中,分子间氢键或范德华力受到破坏,分子内部的相互作用随之减小,而分离纯化有利于晶体结构和三螺旋结构的形成。
目前,有较多的文献报道了关于多糖提取方法对结构影响的研究,但针对分离和纯化工艺是否对多糖结构具有影响的研究相对较少。例如,在多糖脱蛋白的研究中,一般以蛋白质清除率、多糖损失率、多糖得率作为指标,并未深入探讨脱蛋白方法与多糖结构之间的关系。有学者认为,醋酸铅法、三氯乙酸法可能对多糖结构造成破坏,Sevag法、醋酸铅法、三氯乙酸法、氢氧化钠法可能破坏多糖活性。分级纯化法可能会影响多糖结构和活性,金属络合物法可能影响多糖活性。但分离纯化工艺具体如何影响多糖结构的机理机制,进而影响多糖生物活性还鲜有报道。
3.3 化学修饰
多糖分子的修饰是针对性地利用目标基团替代多糖分子上不同的基团(一般为羟基、羧基、氨基)进行化学修饰,使多糖结构发生变化,单糖构成比例、分子质量和形貌特征等也随之改变,从而改善多糖生物活性。目前,常用的化学修饰方法主要有硫酸化、磷酸化、羧甲基化、化学降解修饰和乙酰化等。
硫酸化修饰是利用硫酸基团取代羟基生成多糖硫酸化衍生物的过程。常用的方法有浓硫酸法、氯磺酸-吡啶法和三氧化硫-吡啶法。朱丽丹采用氯磺酸-吡啶法,以离子液体为介质对米糠多糖进行硫酸酯化结构修饰,通过傅里叶红外光谱分析发现,在809.52~836.66 cm处的吸收峰是C-O-S伸缩振动,伸缩振动峰的位置和硫酸根基团在糖环上取代位置有关。氯磺酸-吡啶法对多糖进行硫酸化修饰通常伴随着多糖的降解。但是,在硫酸化修饰凉粉草多糖过程中,多糖分子质量增加,表明硫酸化衍生物的成功制备没有伴随多糖降解,这可能缘于甲酰胺的缓冲作用和硫酸化改性的适度反应条件。Xiao Heng等在利用不同反应条件对海蒿子多糖进行硫酸化修饰后,发现改性导致多糖的单糖组成发生改变,葡萄糖含量显著降低,而其他单糖(岩藻糖、阿拉伯糖、半乳糖、木糖、甘露糖、半乳糖醛酸和葡萄糖醛酸)含量略微增加。
磷酸化修饰是一种常见的结构修饰方法之一,指多糖中的羟基被磷酸基团取代。常用的磷酸化材料有磷酸、三氯氧磷、磷酸氢二钠、磷酸二氢钠和三聚磷酸钠等。何淼等利用多聚磷酸盐法修饰山豆根多糖,红外光谱定性分析发现,1 153.00 cm处的吸收峰为P=O的伸缩振动;1 021.29 cm处为P-O-C的伸缩振动;899.36 cm处为P-O-R的伸缩振动。在三氯氧磷修饰麦冬多糖研究中,张力妮等发现麦冬多糖化学修饰后空间结构发生改变,呈现三螺旋结构,红外光谱研究发现,在1 035 cm和1 120 cm处有P-O-C的伸缩振动吸收峰。
羧甲基化修饰是将多糖分子上引入羧甲基基团,具有成本低、制备简便、试剂易得等特点,是多糖结构修饰常用的一种方法。其原理是将多糖与一氯乙酸在碱性的条件下相互反应,从而将羧甲基引入原多糖残基上的某些羟基上。目前常用的方法有溶媒法和水媒法。吴慧玉采用羧甲基化、硫酸酯化和乙酰化对黄秋葵多糖进行结构修饰,发现经不同方法修饰得到的多糖其分子的不对称性、链构象、分子厚度、分子间距及表面形貌发生改变。
3.4 物理修饰
研究表明,过大的分子质量、复杂的空间结构和高聚合度是阻碍多糖发挥生物活性的主要原因。而对多糖结构采用物理修饰方法,可以达到降低多糖的分子质量、改变多糖空间结构、降低聚合度的目的,有利于增强多糖生物活性。与化学修饰相比,物理修饰具有反应条件温和、环保、操作简便等优点。目前,常用的物理修饰方法有超声波降解修饰、离子辐射修饰和高压均质降解等。
超声波降解修饰具有降解过程和降解程度易控制、易获得目标分子质量、无杂质引入、绿色等优点在多糖结构修饰中得到了广泛应用,其原理主要是利用低频(1 MHz)、高强度(3 W/cm)的超声波通过增加质点振动能量来切断多糖大分子中的糖苷键以降低多糖分子质量,增加水溶性,从而提高一些大分子多糖(如纤维素、壳聚糖、葡聚糖)的生物活性。Yang Haihong等以黑加仑果实为原料,利用超声波处理降解黑加仑果实多糖,发现多糖分子质量随超声时间的延长和功率的增加而减小,使用超声功率300、500 W和700 W超声降解100 min后,黏均分子质量从6.12h10Da分别下降到9.40h10、8.00h10、4.65h10Da,数均分子质量也从8.13h10Da分别降至3.39h10、1.74h10、4.57h10Da。
离子辐射修饰具有反应易控、无需添加物、环保、绿色等优点。其原理是采用γ射线、X射线及电子束等电离辐射诱导多糖发生物质聚合、交联、接枝或降解等物理化学变化以改变多糖的分子结构提高其生物活性。目前,离子辐射修饰在壳聚糖、海藻酸钠、纤维素等多糖降解上都有报道。Henniges等利用γ射线照射木屑后,其纤维素的聚合度发生了降低。
高压均质降解利用高压均质机来实现多糖降解的一种新型纯物理降解方法,具有纯物理作用、产品纯度高、简便、环保等优点。高压均质降解技术在多糖中的应用,目前报道较多的是针对壳聚糖的降解。研究发现,高压均质降解壳聚糖是通过机械作用切断壳聚糖分子链,降低分子质量,降解效果随循环次数的增加而增加,对产物的结构和脱乙酰度基本不改变。Xie Fan等采用高压均质处理(200 MPa、20 min)对紫肉马铃薯多糖进行改性,结果发现其单糖组成发生改变,阿拉伯糖的含量升高,而葡萄糖的含量则降低。Avi等为了研究高压均质化对果胶的影响,以柑橘果胶和苹果果胶为原料进行高压均质化处理,结果表明,高压均质可以使柑橘果胶发生明显解聚,显著降低其分子质量,但对苹果果胶几乎没有影响,这可能与苹果果胶中性糖含量更加丰富、结构更加致密有关。
改变各种植物多糖结构的方法整理见表2。

表2 改变植物多糖结构的主要方法Table 2 Main chemical and physical methods to modify the structure of plant polysaccharides
4 结 语
自然界植物种类繁多,植物多糖结构丰富。植物多糖作为肠道菌群的能量来源,可作为潜在益生元开发差异化的健康产品,具有广阔的应用前景。植物多糖结构复杂,目前构效关系尚未完全明晰,制约了植物多糖精准化利用。随着多糖结构解析的深入,相关研究人员应结合多组学技术,逐渐明确肠道微生物对植物多糖的利用过程;建立具有益生活性的植物多糖快速筛选技术;同时应关注植物多糖与蛋白、脂肪、维生素以及其他功能性成分如何协同调节肠道菌群、促进人体健康。
猜你喜欢
今日农业(2022年14期)2022-09-15
江西水产科技(2022年2期)2022-05-17
大众健康(2021年8期)2021-08-04
科学与财富(2021年36期)2021-05-10
科技视界(2018年1期)2018-03-15
百科知识(2017年10期)2017-05-19
百科知识(2017年3期)2017-03-17
湖北农业科学(2017年1期)2017-03-09
湖北农业科学(2015年16期)2015-10-28
党的生活(黑龙江)(2015年7期)2015-07-14
