
怀玉山三叶青烟草病毒增殖蛋白1基因克隆、亚细胞定位和组织表达分析
2022-06-29 00:40:44洪森荣向琼钰熊晨露徐晨慧徐璐珂陈荣华
浙江农业学报 2022年6期
洪森荣,向琼钰,谢 颖,熊晨露,徐晨慧,徐璐珂,陈荣华,蔡 红
(1.上饶师范学院 生命科学学院,江西 上饶334001; 2.上饶市药食同源植物资源保护与利用重点实验室,江西 上饶 334001; 3.上饶市三叶青保育与利用技术创新中心,江西 上饶 334001; 4.上饶农业技术创新研究院,江西 上饶 334001; 5.上饶市红日农业开发有限公司,江西 上饶334700)
三叶青(Diels et Gilg),为我国特有珍稀濒危药用植物。三叶青可理气健脾,增强免疫,具抗癌、抗炎、解热镇痛、保肝消肿等作用。病毒增殖涉及多种宿主因子与病毒基因组编码因子协同作用。而最近的基因和生化分析已经揭示病毒增殖的一些重要宿主因子,然而关于这些宿主因子认识仍然有限。为了确定烟草病毒高效增殖所必需的宿主因子,有研究分离了拟南芥的两个独立突变体,命名为烟草病毒增殖蛋白1(tobamovirus multiplication 1,TOM1)和烟草病毒增殖蛋白2(tobamovirus multiplication 2,TOM2)。两个独立突变体TOM1和TOM2的烟草病毒增殖降低到低水平,致病突变对烟草病毒的细胞内增殖是隐性且有特殊影响,但对黄瓜花叶病毒或芜菁皱褶病毒(TCV)的细胞内增殖没有影响。因此,克隆三叶青烟草病毒增殖蛋白1基因并进行实时荧光定量PCR(qRT-PCR)分析,对研究烟草病毒增殖蛋白1基因在三叶青中的表达和三叶青烟草花叶病毒的增殖具有重要意义。关于烟草病毒增殖蛋白1研究的报道较少。研究表明,烟草病毒增殖蛋白1(tobamovirus multiplication 1,TOM1)基因被鉴定并发现其编码一种推测的七通道跨膜蛋白。TOM1蛋白与烟草病毒螺旋酶结构域多肽的相互作用以及TOM1及其同源物TOM3同时失活可完全抑制烟草病毒增殖业已被证实。拟南芥TOM1和TOM3基因可编码烟草病毒增殖所需的同源蛋白。尽管拟南芥基因组编码另一个TOM1样基因THH1支持烟草病毒增殖,但作用低于TOM1和TOM3。目前,对三叶青的研究主要集中在化学成分、药理临床、种植栽培、组织培养等方面,关于三叶青烟草病毒增殖蛋白1方面的研究尚未见报道。本文拟利用RT-PCR 技术克隆怀玉山三叶青烟草病毒增殖蛋白1基因,并采用生物信息学方法、烟草叶片瞬时表达和实时荧光定量 PCR进行序列分析、亚细胞定位分析和组织表达分析,为揭示怀玉山三叶青烟草病毒增殖蛋白1的生物学功能提供理论依据,为从分子水平调控怀玉山三叶青烟草病毒增殖提供新思路。
1 材料与方法
1.1 试验材料
怀玉山三叶青2个栽培种怀玉1号和怀玉2号试管苗。
1.2 试验方法
1.2.1 总RNA的提取和cDNA第一链的合成
用Trizol试剂提取怀玉山三叶青怀玉2号试管苗的总 RNA,提取步骤按说明书进行,使用紫外分光光度计和琼脂糖凝胶电泳检测 RNA 的浓度和完整性。以提取获得的 RNA为模板,按照 M-MLV cDNA 第一链合成试剂盒说明书合成 cDNA 第一链。逆转录引物用 Oligo(dT) 18 Primer :5′-GGCCACGCGTCGACTAGTACTTTTTTTTTTTTTTTTTT-3′,具体步骤按说明书进行。
1.2.2 烟草病毒增殖蛋白1(tobamovirus multiplication protein 1,TP1)基因的克隆
利用转录组组装的Unigene序列信息(TRINITY_DN20910_c0_g1),运用 Primer Premier 5.0 设计引物(F:5′-ATGAGGGAGCTGGTCTCCTCC-3′;R:5′-CTAGTTAATAGGGTGATATTGCGCC-3′)。PCR扩增条件:95 ℃ 2 min;95 ℃ 30 s,58.5 ℃ 30 s,72 ℃ 30 s,35个循环;72 ℃ 10 min。PCR 产物经 1%琼脂糖凝胶电泳检测后,将含有目的基因的条带与 pMD19-T 载体连接并用热激法转化到感受态细胞大肠埃希菌()DH5α,经鉴定正确的阳性转化子提取质粒送往上海生工进行测序。
1.2.3 烟草病毒增殖蛋白1基因的生物信息学分析
使用 BioEdit软件翻译基因序列为氨基酸序列,用 ProtParam预测酶的理化性质,用 ProtScale预测酶的疏/亲水性。使用GOR I软件在线预测酶的二级结构。使用SWISS-MOLD在线预测酶的三级结构。采用 WoLFPsort在线预测基因的表达部位。通过软件DNAMAN和 Bioedit进行氨基酸序列比对,利用MEGA5.0软件进行系统进化树的构建。
1.2.4 基因克隆和载体构建
以提供的目的基因序列设计引物(去掉终止密码子TAG/TAA/TGA),CZ-TP1-HⅠ-101F: 5′-TCGACCCCGGGGGTACC(载体同源重组序列)ATGAGGGAAAGGCCTGTGGG-3′;CZ-TP1-HⅠ-101R: 5′-GCCCTTGCTCACCAT(载体同源重组序列)ACGGATGGGATGATACTG-3′,以第一次构建载体为模板,以所述引物进行PCR扩增(采用Vazyme公司phanta max super-fidelity DNA Polymerase)。PCR反应体系(总体积50 μL)包括17 μL ddHO、25 μL 2×Phanta Max Buffer、1 μL dNTP Mix(10 mmol·L)、2 μL模板DNA、2 μL引物1(10 μmol·L)、2 μL引物2(10 μmol·L)和Phanta Max Super-Fidelity DNA Polymease(1 U·μL);PCR反应程序为:95 ℃ 30 s;95 ℃ 15 s,退火温度50 ℃ 15 s,延伸72 ℃ 1 min,39个循环;最后延伸72 ℃ 5 min。将PCR产物进行凝胶电泳。将条带切胶回收,即为目的基因片段。用HⅠ酶切载体pRI101-GFP线性化,回收后和目的基因片段重组反应(采用Thermo公司或TaKaRa公司相应限制性内切酶产品)。酶切反应体系包括4 μL 10×K Buffer、5 μL 载体质粒、2 μLHⅠ和ddHO(补足至40 μL)。酶切产物纯化后与上述PCR产物进行重组反应(重组反应试剂盒为Vazyme公司ClonExpress-Ⅱ One Step Cloning Kit)。重组连接反应体系(总体积10 μL)包括4 μL 线性化载体、1 μL 插入片段、2 μL 5×CE Ⅱ Buffer、1 μL Exnase Ⅱ和ddHO(补足至10 μL)。上述反应液使用移液器轻轻吸打混匀,短暂离心将反应液收集至管底。放置于37 ℃反应30 min,随后立即置于冰上冷却。重组产物转化大肠埃希菌DH5α细胞。挑取PCR阳性的转化子摇菌培养提取质粒,同时扩增产物送测序。扩增和测序引物为插入目的基因两侧的载体序列(目的基因较短则只进行一侧测序),分别为35S-F:5′-GACGCACAATCCCACTATCC-3′;GFP-JR:5′-GGGTGAGCTTGCCGTAGGTG-3′。
1.2.5 烟草叶片亚细胞定位
播种烟草,培养约一个月用于实验;将构建好的载体质粒转入农杆菌GV3101,涂布卡那霉素抗性平板,挑取单克隆于YEB液体培养基在28 ℃摇床内培养2 d,菌体4 000 r·min离心4 min,去上清后菌体用10 mmol·LMgCl(含120 μmol·LAS)悬浮液重悬菌体,调整至0.6左右;用无针头的1 mL注射器吸取农杆菌液,从烟草叶片下表皮(背面)压迫注射;将注射完成的烟草植株弱光培养2 d,即可观察;接种2 d后取注射区域的烟草叶片,制作玻片,在激光共聚焦显微镜(FLUOVIEW FV1Oi,OLYMPUS公司)下观察、拍照;同时以空载体转化的农杆菌作为对照重复上述操作。叶绿体荧光信号说明:叶绿体荧光信号激发波长640 nm,发射波长675 nm;GFP信号说明:绿色荧光蛋白GFP:激发光488 nm,发射光510 nm。
1.2.6 烟草病毒增殖蛋白1基因的组织表达分析
分别取怀玉山三叶青2个栽培种怀玉1号和怀玉2号试管苗的根、茎、叶RNA 500 ng,反转录为cDNA。实时荧光定量 PCR(qRT-PCR,SYBR Green Ⅰ)检测内参基因为。设计引物(F:5′-ACAAGTAAGGTGTAGGCCGAGAA-3′;R:5′-CCACGCAGAAGGTGTTCCA-3′;大小163 bp,退火温度53.1 ℃)。qRT-PCR 检测采用 20 μL反应体系,PCR反应程序:预变性95 ℃ 10 min;变性95 ℃ 10 s,退火延伸60 ℃ 34 s,95 ℃ 15 s,40个循环。使用 2-△△C法计算基因表达水平。实验重复3次。所有数据表示为平均值±标准差,并使用SPSS19.0软件进行统计分析,应用单因素方差分析(One-way ANOVA)检验烟草病毒增殖蛋白1基因组织表达的差异显著性(<0.05)。
2 结果与分析
2.1 怀玉山三叶青烟草病毒增殖蛋白1基因cDNA序列
通过对怀玉山三叶青怀玉1号(HY1组)和怀玉2号(HY2组)块根(同一大棚同一栽培基质同一栽培时间)的转录组分析,再通过差异表达基因分析确定烟草病毒增殖蛋白1基因的核心片段,发现转录组数据库中只有1条1同源基因的序列信息。通过PCR扩增技术(图1),怀玉山三叶青烟草病毒增殖蛋白1基因cDNA总长度为888 bp(图2),G+C 含量为51.58%(图3)。
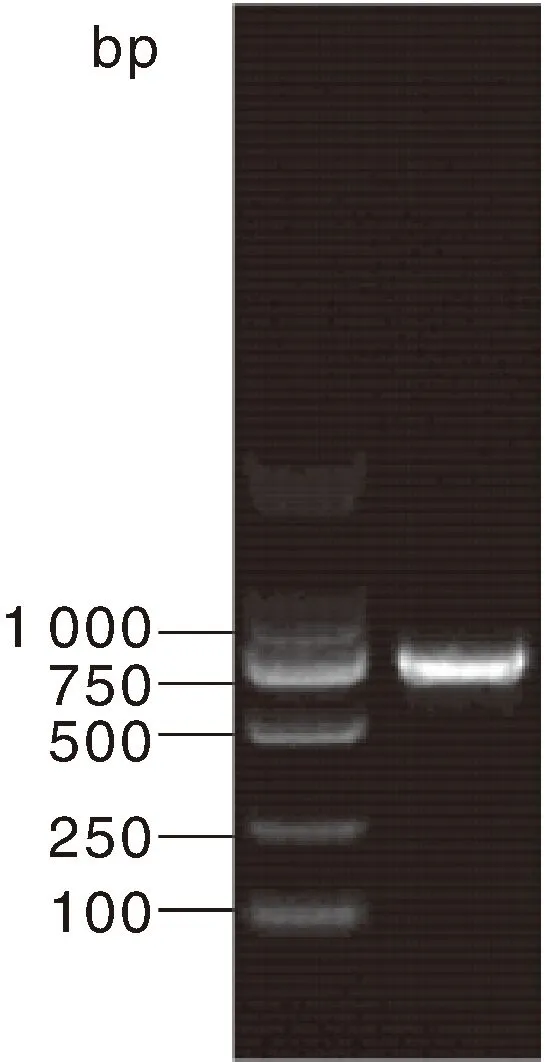
图1 怀玉山三叶青烟草病毒增殖蛋白1基因PCR扩增

图2 怀玉山三叶青烟草病毒增殖蛋白1基因碱基组成
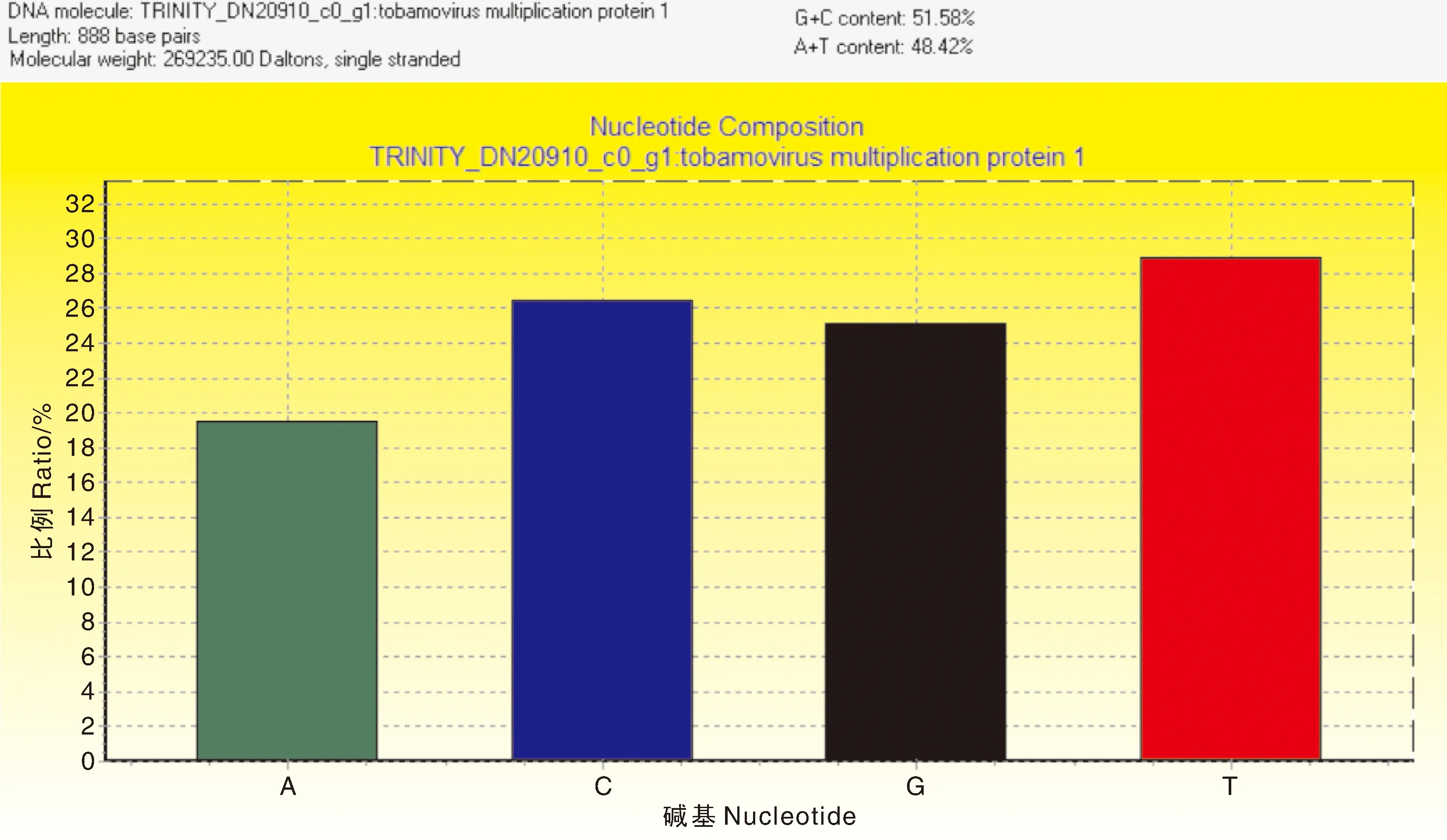
图3 怀玉山三叶青烟草病毒增殖蛋白1基因各碱基的比例
2.2 怀玉山三叶青烟草病毒增殖蛋白1氨基酸序列
Protparam预测显示,怀玉山三叶青烟草病毒增殖蛋白1氨基酸序列见图4。怀玉山三叶青烟草病毒增殖蛋白1由295个氨基酸组成,分子量33 173.36 u,等电点9.16,为疏水性蛋白。各氨基酸的数目和比例为Ala(A)(29,9.8%)、Arg(R)(15,5.1%)、Asn(N)(7,2.4%)、Asp(D)(8,2.7%)、Cys(C)(5,1.7%)、Gln(Q)(8,2.7%)、Glu(E)(11,3.7%)、Gly(G)(13,4.4%)、His(H)(6,2.0%)、Ile(I)(21,7.1%)、Leu(L)(41,13.9%)、Lys(K)(11,3.7%)、Met(M)(3,1.0%)、Phe(F)(20,6.8%)、Pro(P)(11,3.7%)、Ser(S)(19,6.4%)、Thr(T)(12,4.1%)、Trp(W)(6,2.0%)、Tyr(Y)(13,4.4%)、Val(V)(36,12.2%)。带负电荷残基总数(Asp+Glu)为19,正电荷残基总数(Arg+Lys)为26。序列的N端为M(Met),序列的C端为N(Asn)。估计半衰期为:30 h(哺乳动物网织红细胞,体外)>20 h(酵母,体内)>10 h(大肠埃希菌,体内)。不稳定指数:失稳指数(II)计算为37.40,这将蛋白质分类为稳定的。

图4 怀玉山三叶青烟草病毒增殖蛋白1氨基酸序列
2.3 怀玉山三叶青烟草病毒增殖蛋白1亲疏水性分析
从图5可知,高峰值(正值)的区域表示疏水的区域,而负值的“低谷”区域是亲水区域。疏水性结果分析表明,最大疏水值为3.0左右,在该多肽中说明该处的疏水性最强;亲水峰最大值为-2.5左右,整个蛋白质表现出高度的疏水性,说明该蛋白为疏水性蛋白质。
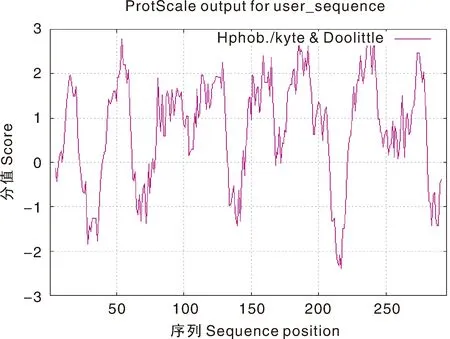
图5 怀玉山三叶青烟草病毒增殖蛋白1亲疏水值分布
2.4 怀玉山三叶青烟草病毒增殖蛋白1二级结构分析
怀玉山三叶青烟草病毒增殖蛋白1的二级结构(图6)预测如下:GOR 预测显示其二级结构由α-螺旋 Alpha helix(Hh,43.73%)、β-片层 Extended strand(Ee,21.69%)、 无规则卷曲 Random coil(Cc,34.58%)构成(图7)。从分布位点上来看,C端和N端含无规则卷曲、β-片层和α-螺旋,且无规则卷曲、β-片层和α-螺旋则散布于整个蛋白质中。
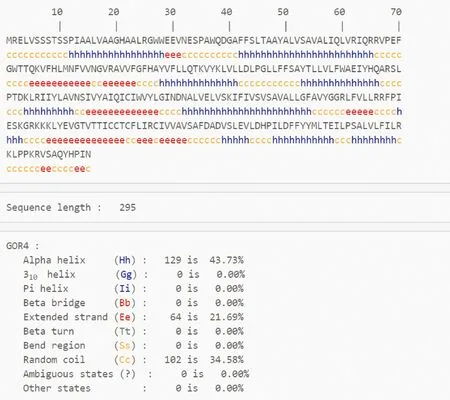
图6 怀玉山三叶青烟草病毒增殖蛋白1二级结构
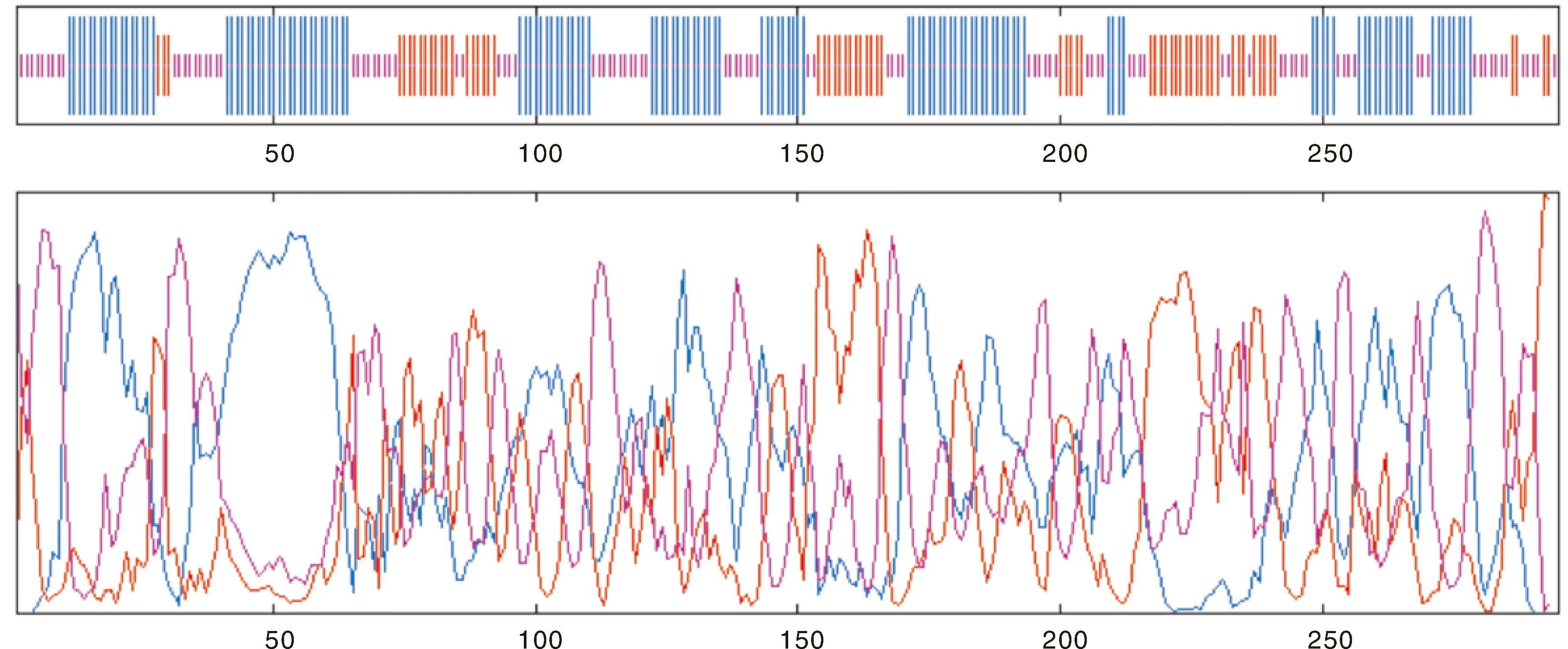
蓝色代表 α-螺旋;红色代表β-片层;紫色代表不规则卷曲。
2.5 怀玉山三叶青烟草病毒增殖蛋白1三级结构分析
SWISS-MODEL预测显示,怀玉山三叶青烟草病毒增殖蛋白1的三级结构为单体(图8)。
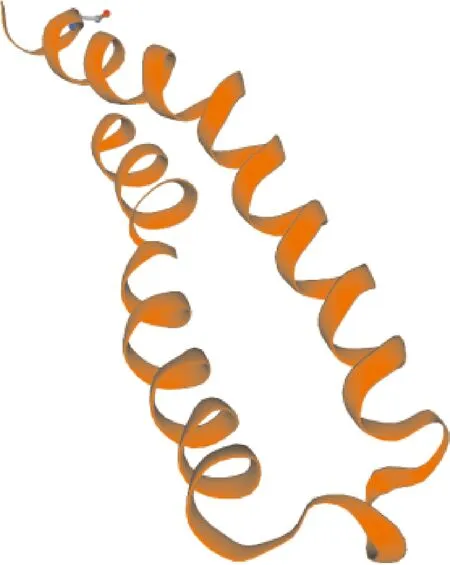
图8 怀玉山三叶青烟草病毒增殖蛋白1三级结构
2.6 怀玉山三叶青烟草病毒增殖蛋白1亚细胞定位预测
采用 WoLFPsort在线软件对怀玉山三叶青烟草病毒增殖蛋白1基因的表达部位进行预测(图9),结果为:定位于内质网中的数量为5.5,内质网-质膜中的数量为4,细胞外的数量为3,细胞质中的数量为2,线粒体中的数量为2,质膜中的数量为1.5。表明怀玉山三叶青烟草病毒增殖蛋白1基因主要存在于内质网、内质网-质膜、细胞外、细胞质、线粒体和质膜中。

E.R.,内质网;E.R._plas,内质网-质膜;extr,细胞外;cyto,细胞质;mito,线粒体;plas,质膜。
2.7 怀玉山三叶青烟草病毒增殖蛋白1系统进化分析
从构建的进化树(图10)中可见,怀玉山三叶青与subsp.(节节麦)、subsp.(硬粒小麦)、(大麦)在一个大分支下,这说明怀玉山三叶青烟草病毒增殖蛋白1在进化上与subsp.(节节麦)、subsp.(硬粒小麦)、(大麦)的亲缘关系较近,尤其是与subsp(节节麦)烟草病毒增殖蛋白1在进化上具有最高的亲缘关系。

图10 怀玉山三叶青烟草病毒增殖蛋白1系统进化分析
2.8 怀玉山三叶青烟草病毒增殖蛋白1同源蛋白的序列比对信息
怀玉山三叶青烟草病毒增殖蛋白1同源蛋白的序列比对信息见图11。图11中※号区域是该蛋白家族的保守结构域。

图11 怀玉山三叶青烟草病毒增殖蛋白1基因氨基酸序列的同源性比较
2.9 怀玉山三叶青烟草病毒增殖蛋白1基因目的片段扩增与瞬时表达载体构建
怀玉山三叶青烟草病毒增殖蛋白1基因目的片段扩增见图12。用HⅠ酶切载体pRI101-GFP线性化,回收后和目的基因片段重组反应。重组产物转化大肠埃希菌DH5α细胞。挑取PCR阳性的转化子摇菌培养提取质粒,对重组质粒进行酶切检测(图13),同时扩增产物送测序。测序比对结果显示:目的基因已插入载体,表达载体构建准确。
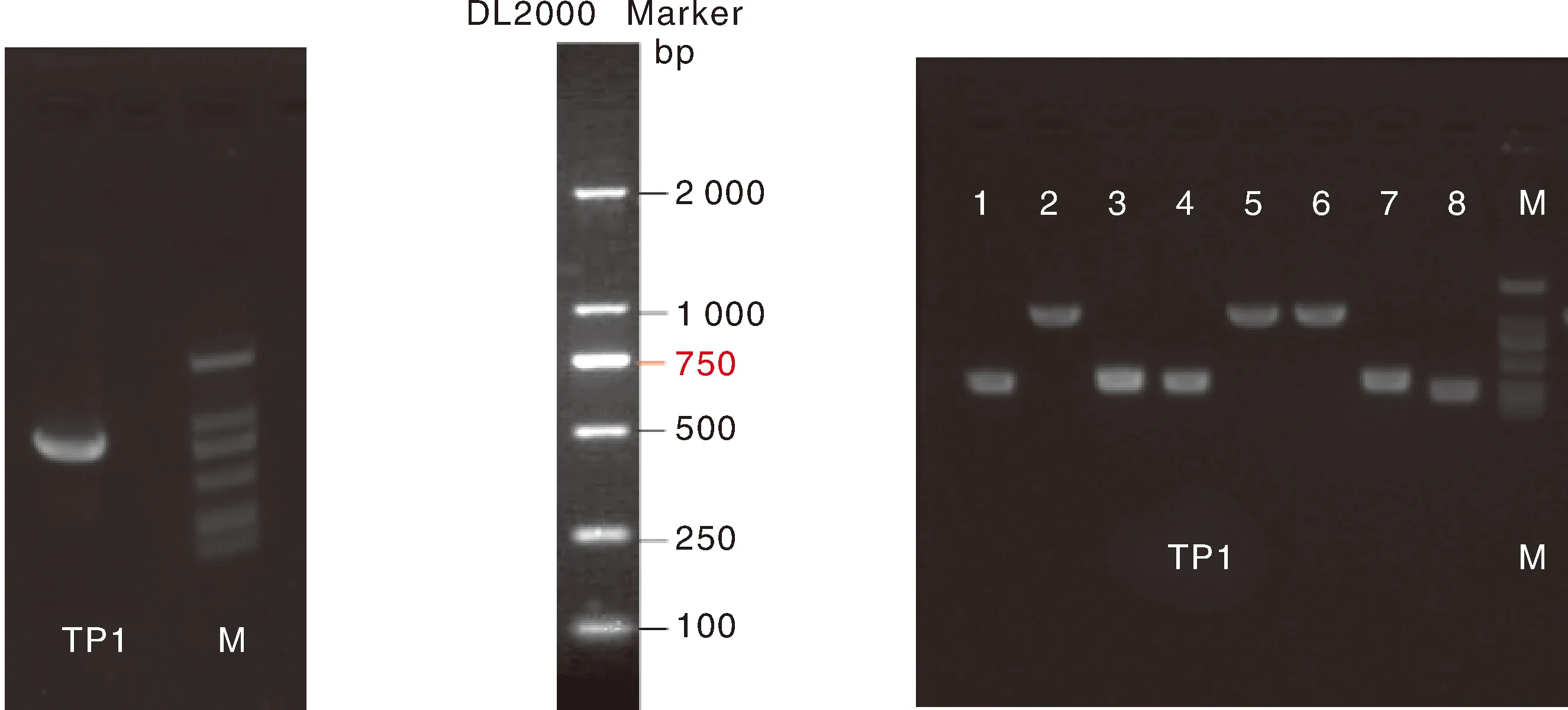
图12 烟草病毒增殖蛋白1基因克隆电泳图(左)和菌落PCR电泳图(右)

第一行是参考序列,第二至三行是构建的GFP载体6号克隆用两端测序引物的测序结果。
2.10 怀玉山三叶青烟草病毒增殖蛋白1亚细胞定位分析
从图14可知,利用激光共聚焦显微镜观察,融合GFP的烟草病毒增殖蛋白1转化烟草叶片只在细胞质(可能包括细胞膜)和细胞核膜观察到绿色荧光,表明烟草病毒增殖蛋白1定位于细胞质(可能包括细胞膜)和细胞核膜。

图14 TP1-GFP基因(上)和GFP对照(下)亚细胞定位照片
2.11 怀玉山三叶青2个栽培种不同器官中烟草病毒增殖蛋白1基因的表达分析
以怀玉山三叶青的为内参,利用实时荧光定量PCR分析怀玉山三叶青烟草病毒增殖蛋白1基因在怀玉山三叶青2个栽培种不同器官中的表达情况,结果显示,怀玉山三叶青烟草病毒增殖蛋白1基因在根、茎、叶中均有表达,但在不同组织器官的表达情况差异显著(图15),其中怀玉2号在叶中表达量最高,怀玉1号在茎中表达量最高。
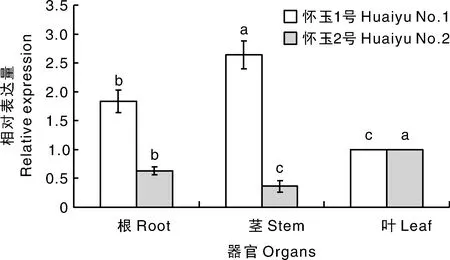
同一品种柱上无相同字母的表示差异显著(P<0.05)。
3 讨论
宿主编码因子在病毒增殖中起着重要作用,与病毒编码因子协同作用。然而,关于这个过程中所涉及的宿主因素的信息是有限的。有研究报道TOM1 mRNA(876 nt)序列由11个外显子组成,编码一个291 aa多肽,该多肽被认为是一种多通道跨膜蛋白。Sos恢复系统(Sos recruitment system,SRS)实验支持TOM1与细胞膜相关的假设,且TOM1与tobamovirus(烟草病毒)编码的复制蛋白的螺旋酶结构域相互作用,TOM1可能作为膜锚参与体内复制复合物的形成。碱基组成和G+C含量基本平衡来看,G+C含量大于 50%,G+C的含量越高,基因稳定性越高,三叶青烟草病毒增殖蛋白1基因1基本处于稳定状态,这可能为三叶青烟草病毒增殖蛋白1基因1稳定遗传与进化提供了保证。从同源性来看,三叶青烟草病毒增殖蛋白1基因1与节节麦、硬粒小麦和大麦同源性较高,说明三叶青烟草病毒增殖蛋白1基因1具有保守性。这些保守区段的发现将为其他物种新的三叶青烟草病毒增殖蛋白1基因1的克隆提供序列依据。
也有研究表明,拟南芥1基因编码一种多倍体跨膜蛋白,是烟草病毒高效增殖所必需的,对1基因具有严重破坏性的突变会使tobamovirus(烟草病毒)的增殖降低到较低水平,但不会完全破坏它。在本实验中,通过烟草叶片亚细胞定位分析表明,烟草病毒增殖蛋白1(TOM1)定位于细胞质(可能包括细胞膜)和细胞核膜中。
在本实验中,实时荧光定量PCR结果显示,烟草病毒增殖蛋白1基因1在怀玉山三叶青2个栽培种中的表达存在器官特异性,怀玉2号在叶中表达量最高,怀玉1号在茎中表达量最高。表明,怀玉2号叶和怀玉1号茎中烟草病毒增殖较快。本研究首次从怀玉山三叶青基因组中克隆出烟草病毒增殖蛋白1基因1的基因,并对克隆的基因序列和其氨基酸序列进行了生物信息学、亚细胞定位和组织表达分析,为三叶青烟草病毒增殖的研究补充了数据和资料。
猜你喜欢
奥秘(创新大赛)(2023年3期)2023-05-06 01:48:20
华人时刊(2022年9期)2022-09-06 01:02:44
红蜻蜓·低年级(2022年5期)2022-05-11 22:04:03
幼儿园(2021年16期)2021-12-06 01:06:36
华人时刊(2020年15期)2020-12-14 08:10:36
红领巾·探索(2020年5期)2020-05-19 15:28:03
家教世界·创新阅读(2020年12期)2020-01-11 01:05:54
浙江中西医结合杂志(2017年2期)2017-01-12 18:23:59
当代化工研究(2016年9期)2016-03-20 16:22:08
广州大学学报(自然科学版)(2015年4期)2015-12-23 11:50:10
