
不同海拔多花黄精各器官碳氮磷化学计量特征
2022-06-28 03:40潘永柱朱强根赵建诚汤郑豪
福建林业科技 2022年2期
潘永柱,朱强根,赵建诚,徐 军,汤郑豪
(1.丽水市林业技术推广总站,浙江 丽水 323000;2.丽水学院生态学院,浙江 丽水 323000;3.浙江省林业科学研究院,浙江 杭州 310023;4.德清县自然资源和规划局,浙江 德清 313200)
生态化学计量学是研究生物体在相互作用过程中多种化学元素平衡关系的一门科学,在反映植物器官的内稳态性、养分利用的高低以及限制性元素判断等方面具有重要作用[1-4]。碳(C)、氮(N)、磷(P)是植物的主要组成元素,其中,C起结构性元素的作用[5],N和P是植物生长必需的营养元素,对植物的生长和生理机能的调节起着重要作用,其生态化学计量特征反映了各元素在不同器官中的分配和相互关系[6],与生境条件共同决定植物生长发育过程和营养水平[7-8]。植物各元素在不同器官间的分配并不一致[9-11],植物的生态化学计量特征在不同年龄、季节、海拔间也存在一定的差异[2,12-13]。目前,生态化学计量的研究多集中于植物的叶片,国内外学者从不同角度对植物叶片的化学计量特征进行了大量研究[14-16],而对于各器官的元素分配及其随海拔变化的研究则鲜有报道。
多花黄精(Polygonatummultiflorum)是百合科黄精属的多年生草本植物,具有益肾、健脾、降血脂、降血糖、抗衰老等作用,药用价值较高,在保健品与化妆品开发领域具有良好的市场潜力[17]。多花黄精多分布在林下、灌丛或山坡阴处,属喜荫植物。海拔梯度对多花黄精具有显著影响,其生长量和多糖含量随海拔升高呈逐渐下降趋势[18]。近年来,围绕海拔梯度上土壤养分组成的分布和变化等研究逐渐增多,海拔梯度变化作为一种自然地理变化,对土壤理化性质、植被分布以及因植被密度不同导致凋落物量的变化均有较大影响,直接或间接地影响土壤碳库大小及碳库组成,从而影响植物的生长发育及植物体内各器官的元素含量分配[19-20]。目前,有关多花黄精的研究主要集中在其生长以及多糖、皂苷、氨基酸、黄酮、矿质元素等成分与含量分析等方面[18,21-23],而对多花黄精各器官C、N、P化学计量特征的研究较少。因此,本研究通过分析不同海拔条件下多花黄精各器官的元素变化规律及其化学计量特征,探讨多花黄精化学计量特征随海拔梯度的协同变化规律,以期揭示不同海拔条件下多花黄精各器官的养分平衡机理及环境适应机制,为多花黄精的合理经营管理提供参考。
1 材料与方法
1.1 试验区概况
试验地位于浙江省松阳县玉岩镇(28°14′—28°36′N、119°10′—119°42′E),属中亚热带季风气候,季风交替明显,温暖湿润,四季分明。该区年平均气温16.6 ℃,年均降水量1700 mm,全年无霜期约236 d,年均日照时间1840 h。土壤为红壤,土层厚度60 cm以上,平均土壤容重1.23 g·cm-3,pH值5.24。
1.2 试验设计
2018年9月,选择4处不同海拔(200、500、800、1100 m)的种植地,取样并测定其土壤养分,各养分含量在不同海拔间无显著差异,土壤温、湿度随海拔升高逐渐降低。2018年10月,对试验地进行林地清理和整地,11月采用带1~2个芽的多花黄精根茎苗栽植,栽植密度4.8万株·hm-2。试验采用随机区组设计,每一海拔梯度内分别设置20 m×20 m的样地3块。
1.3 样品采集与测定
2020年10月,果实基本成熟时,在各样地分别设置3个5 m×5 m的小样方,调查多花黄精的地径、高度;并以平均地径和平均高为参照(标准株),在样地内随机挖取标准株20株,分根、茎、叶、块茎、果5部分取样,分别装袋,并带回实验室。样品经去离子水清洗后于105 ℃杀青30 min,然后85 ℃烘干至恒重,粉碎后过0.25 mm的筛子,装入自封袋保存。
植物样品全C含量采用重铬酸钾外加热法测定[24],全N含量采用高氯酸—硫酸消煮凯氏定氮法测定[24],全P含量采用高氯酸—硫酸消煮钼锑抗比色法测定[24]。
1.4 数据分析
采用Microsoft Excel 2010对试验数据进行统计分析,Origin 8.6进行绘图,SPSS 19.0软件进行单因素方差分析和最小显著性检验。
2 结果与分析
2.1 多花黄精不同器官C、N、P含量特征
由图1~图3可知,叶的C含量在4个海拔梯度间差异不显著(P>0.05);叶的N含量在海拔200 m和800 m显著高于海拔500 m和1100 m(P<0.05);叶的P含量在海拔200 m时显著高于其它海拔(P<0.05)。茎的C含量在海拔200 m和500 m时显著低于海拔800 m和1100 m(P<0.05);茎的N含量随海拔升高逐渐降低,且海拔200 m显著高于其它海拔(P<0.05);茎的P含量,海拔500 m显著高于其它海拔(P<0.05)。果的C含量在4个海拔梯度间差异不显著(P>0.05);果的N含量随海拔升高呈先降低后升高的趋势,海拔200 m和1100 m显著高于海拔500 m和800 m(P<0.05);果的P含量,海拔1100 m显著高于其它海拔(P<0.05)。根的C含量在4个海拔梯度间差异不显著(P>0.05);果的N、P含量,海拔200 m显著高于其它海拔(P<0.05)。块茎的C含量在4个海拔梯度间差异不显著(P>0.05);块茎的N含量,海拔200 m显著高于其它海拔(P<0.05);块茎的P含量随着海拔的升高逐渐降低,且海拔200 m显著高于其它海拔(P<0.05)。

同一器官不同小写字母为不同海拔间差异显著(P<0.05)。下同。图1 多花黄精不同器官的C含量图2 多花黄精不同器官的N含量
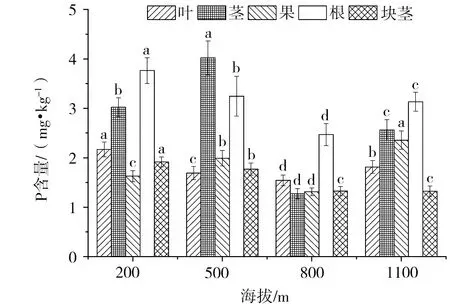
图3 多花黄精不同器官的P含量
2.2 多花黄精不同器官C、N、P化学计量特征
由图4~图6可知,叶的C∶N在海拔200 m与800 m、500 m与1100 m间差异均不显著(P>0.05),且海拔500 m和1100 m均显著高于海拔200 m和800 m(P<0.05);N∶P随海拔的升高呈先降低后升高再降低的趋势;C∶P随海拔的升高呈先升高后降低的趋势,且海拔800 m显著高于其它海拔(P<0.05)。茎的C∶N随海拔升高逐渐升高,且差异显著(P<0.05);N∶P随海拔的升高呈先降低后升高再降低的趋势;C∶P随海拔的升高呈先降低后升高再降低的趋势,且海拔800 m显著高于其它海拔(P<0.05)。果的C∶N随海拔升高呈先升高后降低的趋势,且海拔200 m与1100 m、500 m与800 m间差异均不显著(P>0.05);N∶P、C∶P均随海拔升高呈先降低后升高再降低的趋势,海拔200 m和800 m显著高于海拔500 m与1100 m(P<0.05)。根的C∶N随海拔升高逐渐升高,且海拔200 m显著低于其它海拔(P<0.05);N∶P随海拔升高呈先降低后升高再降低的趋势,海拔200 m显著高于其它海拔(P<0.05);C∶P呈现先升高后降低趋势,且海拔800 m显著高于其它海拔(P<0.05)。块茎的C∶N随海拔的升高呈先升高后降低再升高趋势;N∶P随海拔的升高呈先降低后升高再降低趋势;C∶P随海拔升高逐渐升高,且海拔800 m和1100 m显著高于海拔200 m和500 m(P<0.05)。

图4 多花黄精不同器官的C∶N图5 多花黄精不同器官的N∶P

图6 多花黄精不同器官的C∶P
2.3 多花黄精不同器官C、N、P含量及比值间的相关性
多花黄精不同器官C、N、P含量之间的相关性见表1。块茎C与叶C、叶P、茎N、果C、果N和根C之间均呈显著正相关。块茎N与叶、茎、根的N含量之间均呈极显著正相关,而与果P呈显著负相关。块茎P与茎N、茎P、根N和根P之间均呈极显著正相关,而与叶P、块茎C和块茎N之间均呈显著正相关。

表1 多花黄精不同器官C、N、P含量之间的相关性
多花黄精不同器官C、N、P比值之间的相关性见表2。块茎的C∶N与叶的C∶N呈极显著正相关,与根的C∶N呈显著正相关,与果的N∶P、果的C∶P之间均呈极显著负相关,与叶的N∶P呈显著负相关。块茎的N∶P与叶的N∶P、茎的N∶P、茎的C∶P、果的N∶P、果的C∶P和根的N∶P之间均呈极显著正相关,与叶的C∶N呈极显著负相关。块茎的C∶P与茎的C∶N、茎的C∶P和根的C∶P之间均呈极显著正相关,与叶的C∶P和根的C∶P之间均呈显著正相关。

表2 多花黄精不同器官C、N、P化学计量比之间的相关性
3 讨论与结论
本研究表明,多花黄精各器官的C、N、P含量受外界环境因素的影响较大。海拔变化,植物的生长环境(光照、温度、湿度等)也发生相应的改变,影响植物的生长发育、生理代谢和光合作用等过程,从而影响植物的生物量积累、养分吸收利用等[25-27]。随着海拔升高,太阳辐射能增强,叶片的光合作用更强,叶片内的C含量增加;同时,茎中的C含量随海拔的升高呈增加趋势;而果、根则变化幅度不大;块茎中的C含量随着海拔的升高呈现出下降的趋势。这主要是因为随着海拔升高,温度降低,低温可促进土壤碳的累积,地下细根生物量增加以及树种不同带来较大的生物量输入均可增加碳库累积,与李丹维等[28]的研究结果一致。
N、P含量在各器官中存在显著差异,叶、茎和根的N含量以及叶的P含量均随海拔的升高逐渐降低,这可能是因为高海拔导致土壤温、湿度降低,抑制了根系吸收能力,进而影响各器官的N、P养分含量。已有研究表明,低海拔土壤温度高,土壤风化加快,增加P的积累,土壤中的P含量高,根所能吸收利用的P含量就增加,根中的P元素含量就增加[26]。本研究中,根的P含量随着海拔的升高有显著差异,在海拔200 m处最高,根中P元素的减少能反映出根系对P的选择性吸收随着海拔的升高而减少,分配到其它器官中的P元素含量也减少,导致在块茎中P元素也随着海拔的升高而减少。
块茎是多花黄精收获的主要部分,块茎中的元素含量越高,说明块茎生长越好。本研究中,多花黄精块茎中的C、N、P含量均随着海拔的升高呈下降或先下降后略微升高的趋势,在海拔200 m处最高。研究发现,海拔越高,温度越低,有机物分解和转化速率减慢。低海拔地区土壤中根系吸收的养分元素增多,并运输、分配到块茎中,从而提高块茎中的C、N、P含量。
C、N、P是细胞结构与功能最为重要的生命元素,各元素含量及元素间的化学计量比随植物的生长而发生变化[29]。C∶N可用来衡量植物吸收养分的碳同化能力,在一定程度上反映了植物的营养利用率;而N∶P则用于界定植物生长的养分限制格局[25]。本研究中,随着海拔的升高,改变了土壤温度、湿度等自然环境,多花黄精各器官的C、N、P含量存在差异,C∶N、N∶P等化学计量比也发生了变化。随着海拔的升高,多花黄精各器官的C∶N总体呈升高趋势,说明高海拔的多花黄精具有更高的氮素利用率,这可能与高海拔条件下N的生物有效性和可获得性下降有关[25]。多花黄精各器官N∶P随着海拔的升高呈先下降后升高再降低的趋势,说明不同养分元素对海拔变化响应的敏感性具有显著差异,P元素对海拔升高更为敏感,这与随海拔升高P含量降低的研究结果一致。相关性分析表明,多花黄精各器官间具有复杂的相互作用和协调关系,且受海拔条件及适应策略的影响。
综上所述,多花黄精的叶和茎中的C元素含量高海拔地区比低海拔地区高,而N含量低海拔地区高于高海拔地区。多花黄精根与块茎中的C、N、P含量均表现为低海拔高于高海拔,在低海拔多花黄精的地下部分生长状况比高海拔好。本研究结果表明:在海拔200 m处多花黄精块茎中元素含量高,其它器官的元素含量也较高,因此,在海拔200 m处种植多花黄精将得到更高的品质与产量。同时,在低海拔地区施肥应考虑N、P均衡,高海拔地区施肥则需增施磷肥。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
新农业(2022年16期)2022-11-07
军事文摘(2022年14期)2022-08-26
中国种业(2022年7期)2022-07-21
散文诗(2021年22期)2022-01-12
婚育与健康(2020年12期)2020-02-26
环球时报(2019-05-23)2019-05-23
家庭医药·快乐养生(2015年8期)2015-09-10
现代农业研究(2009年3期)2009-04-14
饮食科学(2009年3期)2009-03-23
