
基于Micro-CT的玉米自交系茎秆维管束表型高通量获取与精准解析
2022-06-24 04:02:16赵欢王璟璐张颖廖生进杜建军郭新宇赵春江
江苏农业科学 2022年12期
关键词:区带
赵欢 王璟璐 张颖 廖生进 杜建军 郭新宇 赵春江
摘要:针对玉米自交系茎秆不同节位形态特征差异大、维管束分布不同等特点,利用Micro-CT扫描技术及基于区带的维管束表型解析方法,对茎秆基部第3节、穗位节、顶位节节间进行显微图像获取,实现茎秆不同节间横切面、表皮区、周皮区、髓区、维管束相关表型的精准解析。研究结果表明,基于区带的维管束表型解析方法适用于茎秆不同节间维管束表型解析,可获得表征茎秆节间组织和维管束形态、几何性状、分布等共47项指标,获取效率高达20 s/张图像。同传统方法相比,基于区带的维管束表型解析方法效率高、通量大,且可获取的表型指标类型多。描述性统计分析结果显示,同茎秆横切面长轴长等传统指标相比,基于区带的维管束表型解析方法获取的功能区表型指标变异系数较大,能更好地反映出不同节间、不同自交系间的差异;且在不同自交系间,基部第3节的周皮区维管束数量(P=7.79×10-29)和穗位节的周皮区维管束面积占比(P=1.79×10-26)差异最显著。此外,从基部到顶部,表征横切面及各功能区几何、形态性状以及维管束数量、面积相关的表型指标呈依次减小的趋势;表征维管束分布的表型指标呈依次增加的趋势;功能区面积占比分布趋势不同,其中周皮区面积占比在基部节间最大,髓区面积占比在穗位节最大,表皮区面积占比在顶位节最大。本研究实现了玉米自交系茎秆不同节间维管束表型的高通量获取与精准解析,初步揭示了表型指标在不同节间的分布趋势,可为后期茎秆表型遗传解析和性状定向改良提供参考。
关键词:玉米茎秆;Micro-CT;区带;维管束表型;高通量获取;精准解析
中图分类号:S126;S513.01 文献标志码: A
文章编号:1002-1302(2022)12-0201-11
收稿日期:2021-12-26
基金项目:国家自然科学基金(编号:31801254);北京市农林科学院作物表型协同创新中心项目(编号:KJCX201917)。
作者简介:赵 欢(1997—),女,四川广元人,硕士研究生,主要从事作物表型组学研究。E-mail:3321621249@qq.com。
通信作者:赵春江,博士,研究员,中国工程院院士,主要从事农业信息化关键技术研究。E-mail:zhaocj@nercita.org.cn。
随着高通量测序技术和信息技术的飞速发展,作物科学研究进入了大数据、大平台、大发现的新时代[1-3]。近年来,以高通量、自动化、智能化为主要特征的表型组学技术及平台系统层出不穷,使得多尺度、高精度、自动化的表型检测成为可能,为实现作物多维度表型精准鉴定奠定了良好基础。在此背景下,基于组学的茎秆维管束形态、分布、关联性等高通量、精准性、系统性研究对助力作物茎秆维管束的表型特征分析和遗传解析具有重要意义[1-2]。
玉米茎秆维管束是“流”的重要组成部分,由木质部和韧皮部构成,负责水分和有机养分的运输。由于受限于显微表型检测设备分辨率和表型解析手段,玉米茎秆维管束研究进程缓慢。传统玉米茎秆维管束表型检测主要依赖显微镜进行局部观察。显微镜虽然分辨率高,但成像视野小,仅能获取预先选择的、特定组织中的维管束数量、面积等表型指标,且无论是人工测量还是应用商业软件,都需要大量人工参与,费时费力[4-7]。因此,基于显微镜的观察方法无法满足大规模、系统性的玉米茎秆维管束表型特征分析需求,迫切需要找到一种高通量、自动化的维管束表型研究方法来打破困局。2014年,Legland等利用BlueBox宏观成像系统对人为去除表皮和周皮的玉米茎秆横切面进行成像分析,获得了维管束空间分布相关性状[8]。2015年,Heckwolf等通过平板扫描仪获得玉米茎秆整个横切面图像,提出一种自定义图像处理程序对图像进行自动化解析,成功实现在样品无损的情况下对髓组织内部维管束表型指标的提取,但并未实现对周皮维管束的表型获取[9]。2017年,Legland等基于玻片扫描仪对Fasga染色后的整个茎秆横切面进行成像,量化各组织区块成色单元,为茎秆外围维管束相关表型测量提供了可能[10]。
鉴于传统成像系统及表型提取手段的诸多不足,通过二维成像技术提取玉米茎秆维管束表型仍存在较大困难。如今,显微计算机χ射线断层扫描(Micro-CT)等三维成像技术在作物表型研究领域得到越来越多的关注[11-15],为玉米维管束性状的提取提供了新的技术途径。Micro-CT系统作为一种高分辨率、高精度仪器,不仅能对植株内部显微表型进行精准获取,还能最大程度地保留样品的完整性。基于Micro-CT扫描技术,可以精准定量玉米茎秆在伸长期和抽雄期的显微指标,实现不同生长阶段的实时观测[16];还可以获取不同品种玉米茎秆不同节间的解剖特征,进行解剖表型与抗倒伏性的关系研究[17]。基于CT图像,Zhang等研发出专用于维管束表型分析的图像处理软件VesselParser 4.0,该软件根据茎秆内物质分布(CT值),将茎秆横切面精准划分为表皮区、周皮区和髓区,并提出了基于区带的表型解析方法。该方法的提出与应用不仅实现了茎秆横切面的组织划分,还实现了维管束分类以及维管束数量、面积和空间分布等显微表型特征的自动化获取[18]。目前,基于区带的维管束表型解析方法已成功应用于玉米茎秆基部第3节维管束的表型研究及遗传解析,而尚无在茎秆其他节间的应用报道。
本研究针对玉米自交系茎秆不同节间形态特征差异大、维管束分布不同等特点,利用Micro-CT扫描技术和基于区带的维管束解析方法,对玉米自交系茎秆基部第3節、穗位节和顶位节进行维管束表型高通量获取及精准解析,并基于获取的茎秆横切面及各区带几何形态性状和维管束数量、面积及分布等47项表型指标,开展玉米不同节间的差异分析,以期为进一步的遗传学研究和性状定向改良提供参考。
1 材料与方法
1.1 供试材料4FC70124-2524-41CF-BEF5-849E79D3D36F
试验材料为玉米关联分析群体[19]中茎秆粗细不同的36个自交系,材料来源地涵盖了热带、亚热带及温带等多气候带。具体材料信息如下,非坚秆综合种(non-stiff stalk,NSS):MO113、GEMS58、By4839、M153、By813、BEM、Dan360、Zheng28、P138;坚秆综合种(Stiff Stalk,SS):GEMS14、832、Tie7922、U8112、GEMS52、B110、Zheng32、GEMS13、GEMS4;热带种质材料(Tropical-subtropical,TST):GEMS31、CIMBL55、CIMBL50、CIMBL69、CIMBL63、CIMBL90、CML69、CML170、CML171;混合材料(Mixed):SK、CIMBL154、Ye515、Zheng22、TY2、TY7、Ye107、Zhong69、3H-2。
1.2 田间试验设计及样品制备
供试材料于2018年4月28日在北京市农林科学院通州试验基地(116°E,39.69°N)种植,采用完全随机区组设计,8次重复,每个自交系种植4行,行长2.1 m,行距0.6 m。种植密度和水肥管理均参照当地大田生产模式。
在玉米吐丝期(种植73 d后)选取长势一致的植株进行取样,每个小区取3株作为重复。获取各自交系基部第3节、穗位节(最大果穗着生处)和顶位节的节间中部切片,获取长度2 cm以上,取样后及时放置在甲醛-乙酸-乙醇(FAA)固定液(70%乙醇、100%甲醛、100%乙酸体积比为90 ∶5 ∶5)中进行固定。
1.3 茎秆样本前处理及CT扫描
(1)样本脱水。样本固定完成后,从固定液中取出,进行系列脱水处理,具体流程为:70%乙醇 4 h→100%乙醇 4 h→100%乙醇 4 h→100%叔丁醇 4 h→100%叔丁醇 4 h。(2)樣本超低温冷冻。叔丁醇代替完成后,将样品从叔丁醇中取出,放置 -80 ℃ 超低温箱中冷冻24 h以上。(3)超低温冷冻干燥。将经超低温冷冻的样品放置于超低温冷冻干燥仪(LGJ-10E,中国)的样品仓中,-30 ℃ 干燥3~4 h。(4)固态碘染色。将干燥后的样品加固态碘进行染色,每个样品中大致放3~5颗固态碘珠,待染色均匀后进行CT扫描。(5)CT扫描。使用Skyscan 1172 X射线计算机断层摄影系统(Bruker Corporation)进行样品扫描,扫描参数设置参照Zhang等设置的CT扫描参数[17]。(6)图像重构。利用Skyscan NRecon软件(Bruker Corporation)进行重构,亨氏单位(HU)统一设置为-500~9 40,获得一系列玉米穗位节横截面为8位(8-bit)的图像文件格式(bmp),用于后期表型特征提取与分析。(7)表型指标提取。使用VesselParser 4.0对茎秆CT图像进行处理,自动获取表型指标。
1.4 表型数据分析
利用R语言(https://cran.r-project.org/)进行数据统计分析。首先,采用“lm”函数进行线性回归分析,建立实测值与VesselParser 4.0软件提取值之间的线性模型;然后,利用psych包对表型数据进行描述性统计分析,结合CT图像进行茎秆不同节间形态及表型指标对比分析;最后,对表型指标进行不同自交系间或不同节间的单因素方差分析。数据分析结果的可视化由ggplot2包呈现。
2 结果与分析
2.1 不同节间显微成像及基于区带的维管束表型特征解析
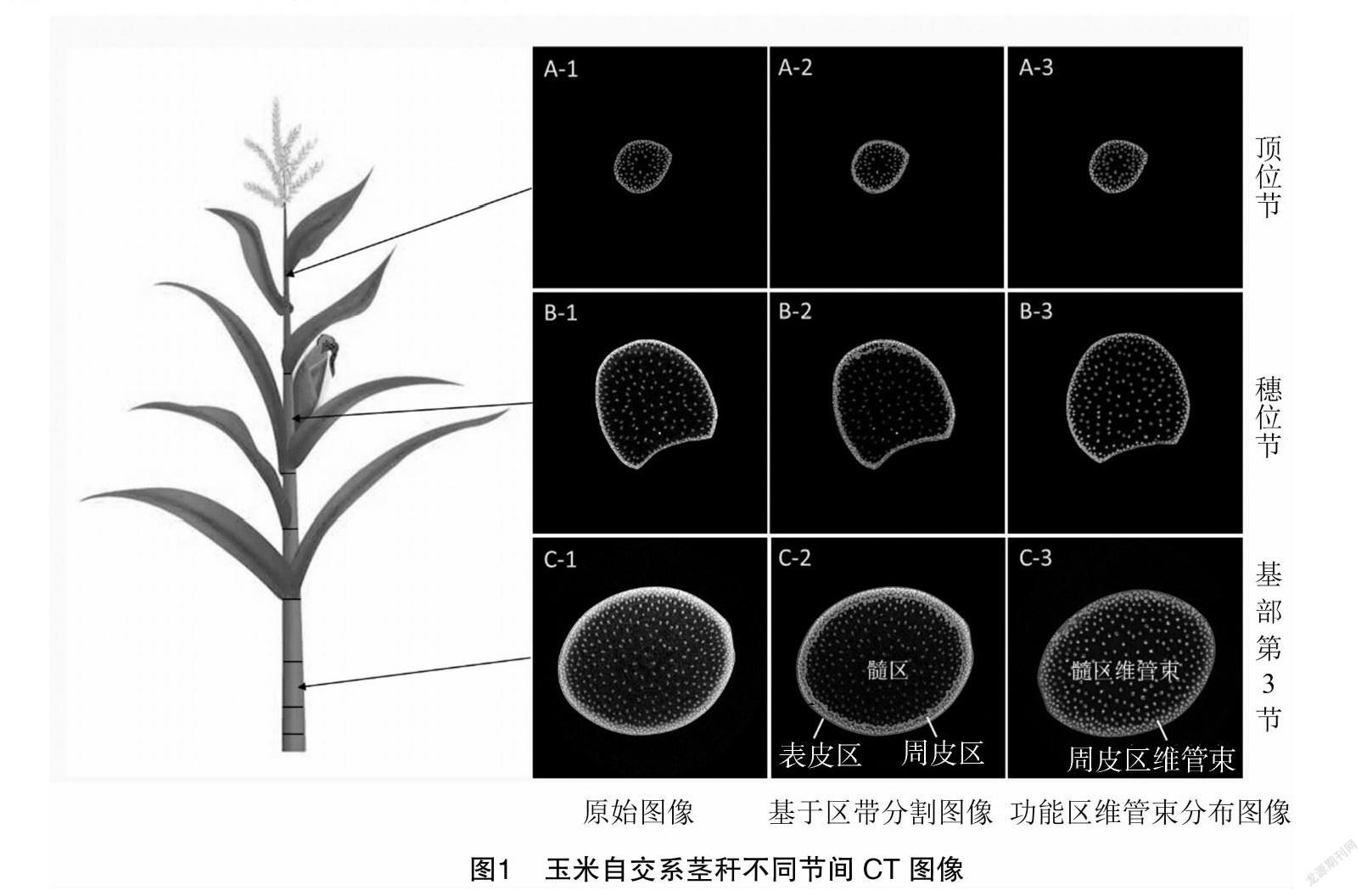
参照Zhang等已有的基部第3节维管束表型解析结果[18],利用Micro-CT扫描技术同时对玉米茎秆上中下关键的基部第3节、穗位节、顶位节进行显微图像获取。从图1中原始图像可以看出,在外部形态上,从基部第3节到穗位节再到顶位节,玉米横切面呈椭圆—扇形—圆形变化,横切面面积依次减小。维管束分布在周皮和髓组织,其中周皮维管束较大,分布密集,髓组织维管束较小,分布松散。在基部第3节中,周皮维管束有3~4层,维管束间分布紧密;穗位节的周皮维管束有1~2层,但分布不均匀,其中凹面分布着1层维管束,凸面分布着2层维管束;在顶位节中,周皮分布着1层间距稀疏的维管束。
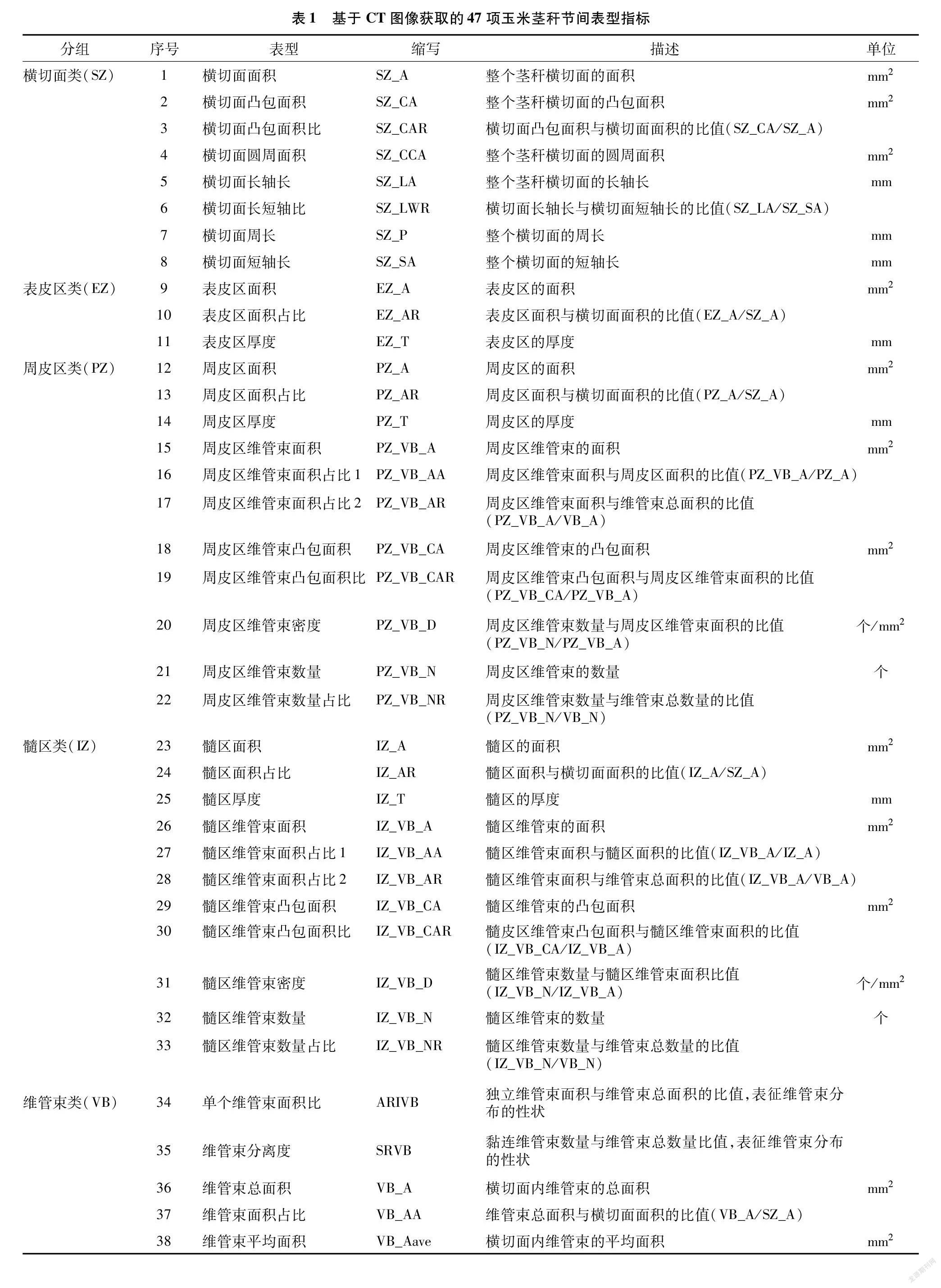
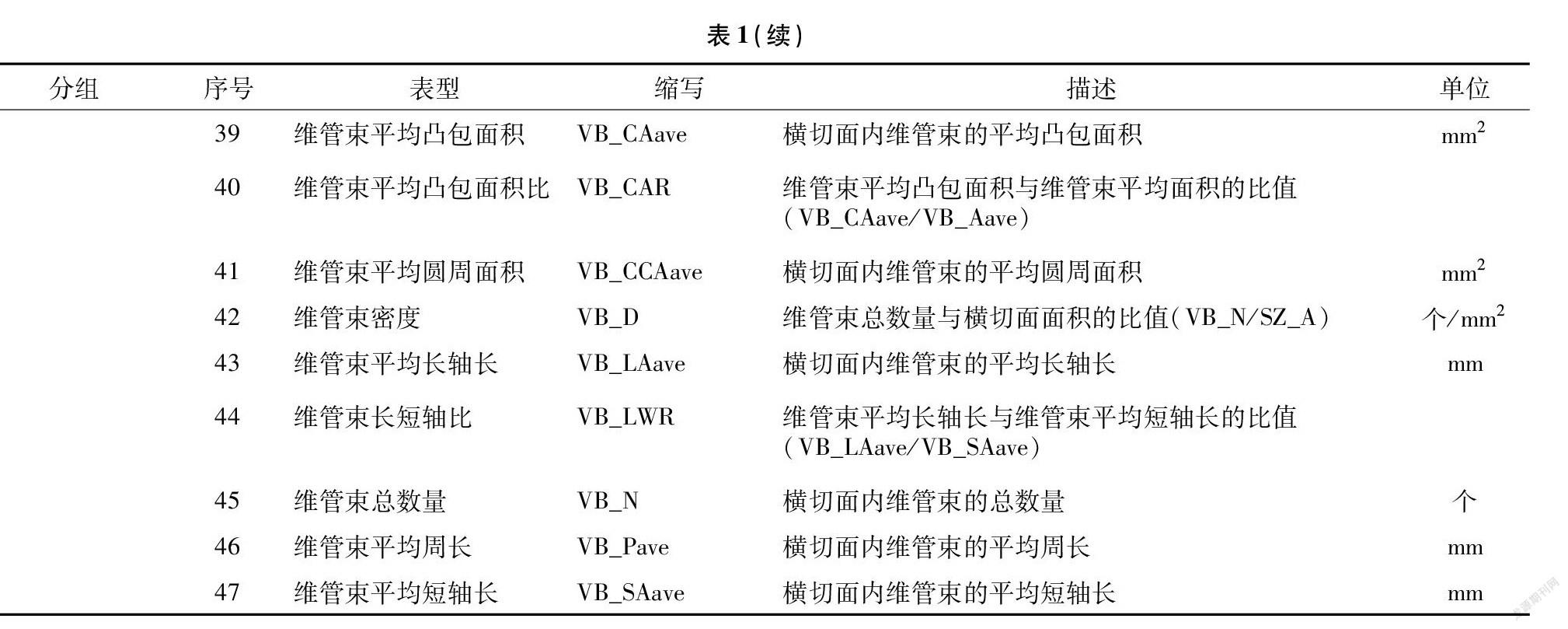
采用基于区带的维管束表型解析方法对不同节间显微图像进行解析,可将茎秆横切面分割为3个功能区,分别为表皮区、周皮区和髓区。结果表明,针对基部第3节内部物质分布(CT值)提出的基于区带的维管束表型解析新方法不受茎秆外部形态和内部解剖结构的影响,在茎秆穗位节和顶位节间表型解析均具有较好的适用性。对玉米茎秆不同节间横切面进行功能区划分后,维管束根据分布位置差异,分为周皮区维管束和髓区维管束。按照传统维管束分类标准[20],即根据维管束面积和结构将其分为大维管束和小维管束,本研究使用的基于区带的维管束表型解析方法除维管束面积外,还参考了维管束内部空腔大小、CT值和分布密度,因而能够反映茎秆不同节间内部物质含量、维管束结构特征和分布变化。利用该方法处理单张CT图像,可一次性自动获取47项表型指标(表1),可分为茎秆横切面类(8项)、表皮区类(3项)、周皮区类(10项)、髓区类(10项)和维管束类(13项)。其中横切面类、维管束类归类于传统表型指标,表皮区类、周皮区类、髓区类表型指标归类于功能区表型指标。
为评价采用VesselParser 4.0获取的穗位节和顶位节表型指标精度,利用ImageJ软件手动统计穗位节和顶位节节间的维管束总数量(VB_N),然后与VesselParser 4.0自动解析的VB_N进行线性回归分析,结果如图2所示。在穗位节中,线性回归模型的决定系数r2为0.98;在顶位节中,线性回归模型的决定系数r2为0.95。两节位的模型决定系数均接近 表明基于VesselParser 4.0软件获取的 VB_N 与实测值吻合程度高,说明本研究采用VesselParser 4.0获取的茎秆节间表型指标精度高,可用于后续不同节间的特征分析。4FC70124-2524-41CF-BEF5-849E79D3D36F
由表2可知,同石蜡切片与人工测量等传统维管束表型获取方法相比[21-22],Micro-CT扫描技术简化了前期制样流程,基于区带的维管束表型解析方法可在 0 s 左右完成单张图像的表型解析,且支持批处理,大大提高了茎秆维管束表型的获取效率,适合大群体维管束表型指标高通量获取。此外,传统方法获取的维管束表型指标(如大、小维管束数量和面积)是在显微图像的局部区域中获取;而本研究是基于完整的横切面图像提取到表征茎秆横切面及各区带几何、形态性状,以及维管束数量、形状、面积、分布等几大类重要表型指标,获取的表型指标类型丰富,且数量多达传统研究方法的5倍以上。以上结果表明,基于区带的维管束表型解析方法不仅能实现玉米不同节位茎秆维管束表型指标的高通量获取,还能实现表型指标的精准解析。
2.2 玉米茎秆不同节间表型指标描述性统计分析
分别对玉米茎秆基部第3节、穗位节、顶位节的47项表型指标进行描述性统计,进而分析不同节间表型特征。以表征玉米茎粗的表型指标SZ_LA作为参照,对比分析各项表型指标的变异情况。
由表3可知,基部第3节的SZ_LA变异系数(CV)为16.35%,其余指标中有26项表型指标的变异系数大于16.35%,其中功能区表型指标占一半以上(表皮区3项,周皮区6项,髓区6项)。在基部第3节中,变异系数最大的表型指标为IZ_VB_CA(CV=42.25%),是SZ_LA的2.6倍左右。在穗位节中,有28项表型指标的变异系数大于SZ_LA的变异系数(CV=15.42%),其中功能区表型指标有16项(表皮区3项,周皮区7项,髓区6项)。穗位节的VB_LWR变异系数(CV=33.75%)最大,是 SZ_LA 的变异系数的2.2倍左右。顶位节的 SZ_LA 變异系数为19.71%,其余指标中有25项表型指标的变异系数大于19.71%,其中表皮区3项,周皮区5项,髓区6项。而且IZ_VB_CA在顶位节中变异系数(CV=43.36%)最大,是SZ_LA变异系数的2.2倍。
描述统计结果显示,SZ_LA在不同节间的变异系数不同,尤其是在穗位节时最小,这可能与穗位节外部形态结构相关。此外,以SZ_LA为变异系数分界线,高于该分界线的表型指标中,至少有一半指标为功能区表型指标,特别是基部第3节和顶位节表型变异系数最大的表型指标均来自功能区类(IZ_VB_CA)。结果表明,同传统指标相比,基于区带的维管束表型解析方法获取的功能区表型指标能更好地反映出不同自交系间的差异。
2.3 玉米茎秆不同节间表型指标方差分析
分别对基部第3节、穗位节、顶位节的47项表型指标进行不同自交系间的差异分析。方差分析结果(表4)显示,基部第3节的47项表型指标在不同自交系间均存在显著差异(P<0.05),其中 SZ_LA 在不同自交系间的P=5.11×10-18,与显著性最高的PZ_VB_N(P=7.79×10-29)相差11个数量级。在穗位节中,47项表型指标均在不同自交系间存在显著差异(P<0.05)。SZ_LA在不同自交系间的显著性为P=5.96×10-2 与显著性最高的 PZ_VB_AR(P=1.79×10-26)相差5个数量级。在顶位节中,除SRVB外,其余46项表型指标在不同自交系间均存在显著差异(P<0.05)。其中,差异性最高的表型指标为VB_N(P=1.25×10-21),与 SZ_LA(P=8.20×10-14)相差7个数量级。
方差分析结果还显示,尽管基部第3节、穗位节和顶位节的47项表型指标大多在P<0.05水平上存在显著差异,但是不同节间差异最显著的表型指标却各不相同。其中,基部第3节和穗位节差异最显著的指标均为功能区的周皮区类表型指标,进一步表明基于区带进行维管束解析更有助于茎秆维管束表型精准鉴定。此外,同描述统计结果相似,穗位节的SZ_LA在不同自交系间的差异显著性水平高于其他节间,可能与穗位节外部形态结构相关。
2.4 玉米不同节间维管束表型指标随叶序分布趋势研究
为研究表型指标在单株尺度内的分布情况,对47项表型指标做关于不同节间的方差分析。结果显示,除SZ_LWR外,其余46项表型指标在基部第3节、穗位节和顶位节间均存在显著差异(P<0.05)。随后,对差异显著的46项表型指标进行3个节间的多重比较分析,进一步探究各表型指标在基部第3节、穗位节、顶位节的具体分布,并将结果进行数据可视化。
由图3可知,除茎秆横切面类表型指标和维管束类表型指标等传统表型指标,基于区带获取的新表型指标在3个节间具有明显的分布规律。首先是横切面类(SZ_A、SZ_CA、SZ_CCA、SZ_P、SZ_LA、SZ_SA)和维管束类(VB_N、VB_A、VB_Aave、VB_CAave、VB_CCAave、VB_LAave、VB_SAave、VB_Pave、ARIVB)等传统表型指标,从基部第3节到穗位节再到顶位节呈依次减小的趋势,并且两两节间差异显著。其次是基于区带获得的新表型指标,功能区面积(EZ_A、IZ_A、PZ_A)、功能区厚度(IZ_T、PZ_T)以及功能区维管束数量(IZ_VB_N、PZ_VB_N)、面积(IZ_VB_A、PZ_VB_A)和凸包面积(IZ_VB_CA、PZ_VB_CA),从基部到顶部呈依次减小的趋势,并且两两节间差异显著。
同VB_D分布一样,PZ_VB_D和IZ_VB_D从基部第3节到穗位节再到顶位节呈依次增加趋势。此外,表型指标EZ_AR、PZ_VB_AA、VB_LWR也呈现同样的分布趋势,并且两两节间差异显著。
此外,IZ_AR、IZ_VB_CAR从基部第3节到穗位节再到顶位节呈现先增大后减小的趋势,且在穗位节时显著大于其他2个节间。而PZ_AR、SZ_CAR呈现相反的分布趋势,且在穗位节时显著小于基部第3节和顶位节。4FC70124-2524-41CF-BEF5-849E79D3D36F
综合差异分析结果可知,47项表型指标中有36项在基部第3节、穗位节和顶位节具有明显的分布趋势,其中有18项表型指标来自各功能区。此外,从基部到顶部分布趋势并非依次减小的表型指标主要来自各功能区,如IZ_AR、PZ_AR,这些表型指标可为后期进行节间功能研究和性状定向改良提供参考。
3 讨论与结论
3.1 基于区带的玉米不同节间维管束表型特征精准解析
基于区带的维管束表型解析方法于2021年由Zhang等首次提出[18]。该团队通过Micro-CT扫描技术和图像处理软件VesselParser 4.0对玉米基部第3节的维管束表型进行研究,并根据茎秆内物质分布将横切面分割为3个功能区,实现了对玉米茎秆基部第3节横切面的精准分割以及維管束结构及分布变化的特征提取。本研究基于Zhang等的研究结果[18],首次将基于区带的维管束表型解析新方法应用于穗位节和顶位节,获取效率高达20 s/张图像,相较传统手段一次性可获得的表型指标增至5倍以上,实现了表型指标的高通量获取。最终在3个节间均获取了包括茎秆横切面及各区带几何形态性状和维管束数量、面积及分布等在内的47项表型指标。对比分析不同节间CT图像,结果表明,虽然穗位节和顶位节在外部形态和内部解剖结构上与基部第3节存在明显差异,但是应用于基部第3节横切面的功能区分割方法同样适用于穗位节和顶位节的研究,表明基于区带的维管束表型解析方法对玉米茎秆不同节位均具有较好的适用性。此外,穗位节和顶位节的VB_N线性回归模型的决定系数均高于0.95,表明通过VesselParser 4.0软件自动解析的表型指标精度较高。
3.2 玉米不同节间维管束表型特征差异分析
玉米茎秆不同节间维管束表型特征不同。47项表型指标在不同自交系间的差异分析结果表明,尽管多项表型指标在基部第3节、穗位节、顶位节间存在显著差异,但是不同节间差异最显著的表型指标却并不相同。其中,不同节间维管束表型差异最显著的表型指标分别为基部第3节的PZ_VB_N(P=7.79×10-29)、穗位节的PZ_VB_AR(P=1.79×10-26)和顶位节的VB_N(P=1.25×10-21),其中前2项指标均为功能区表型指标,进一步表明基于区带进行维管束解析更有助于茎秆维管束表型精准鉴定。
已有研究表明,基部第3节是植株抗倒伏研究的重要对象[23]。蒋傲男等的研究表明,维管束数量与基部第3节穿刺强度呈显著负相关[24];李波等研究发现,小维管束数量在茎秆稳定性方面发挥重要作用[25]。在本研究的方差分析结果中,基部第3节的PZ_VB_N和VB_N在不同自交系间差异最显著,据此推断这2项表型指标可能与玉米茎秆抗倒伏性的相关程度较高。在穗位节相关的研究中,何启平等研究发现由茎节维管束等横向分支组成的穗部维管束系统是养分从植株运输到籽粒中的重要保障[26]。Ren等研究发现,穗位节间大维管束面积占总维管束面积比例的均值小于小维管束面积占总维管束面积的比例,而增施氮肥有利于增加小维管束面积占总维管束面积的比例,提高物质运输速率,加快光合产物向库运输[5]。侯彦龙等也指出维管束面积大有利于营养物质运输[27]。而本研究在穗位节间表型指标关于不同自交系间的差异分析结果表明,PZ_VB_AR、IZ_VB_AR、IZ_VB_N在不同自交系间差异最显著。相较前人的研究,本研究获取的表型指标更精细,分区特征更明显,可为穗位节的物质运输相关研究提供参考。在玉米茎秆上部节间维管束的研究中,王越等利用198份玉米自交系对雄穗中部维管束性状开展不同杂种优势群的比较,结果发现大、小维管束数量和单个大、小维管束面积在不同自交系间的差异均达到极显著水平[28]。杨硕等利用172份玉米自交系对茎秆上部维管束数量进行差异分析,结果同样表明大、小维管束数量在不同自交系间呈极显著差异[7]。而在本研究中,不同自交系在顶位节的差异分析结果显示,除顶位节的VB_N、IZ_VB_N外,VB_AR也在0.05水平上存在显著差异。
3.3 玉米不同节间维管束表型指标随叶序分布趋势
茎秆维管束表型指标在单株尺度内茎秆不同节间分布趋势明显。对获取的47项表型指标进行不同节间分布趋势研究发现,表征横切面及各功能区几何、形态,以及维管束数量、面积相关的表型指标从基部到顶部呈依次减小的趋势。这一结果与前人的研究结果一致,如Ren等指出大、小维管束数量从基部节、穗位节、穗柄到穗轴呈减小趋势[5];赵雪等指出玉米茎秆维管束数量从基部向上逐渐减小[29]。而表征维管束分布的表型指标从基部到顶部呈依次增加的趋势。此外,本研究发现功能区面积占比从基部第3节到穗位节再到顶位节的分布趋势不同,其中EZ_AR呈依次增加的趋势,IZ_AR呈先增加后减小的趋势,PZ_AR呈先减小后增加的趋势。
综上所述,本研究利用Micro-CT扫描技术及基于区带的维管束表型解析方法,实现了玉米自交系不同节间维管束表型的高通量获取与精准解析,并首次证明基于区带的维管束表型解析方法对不同节位均具有较好的适用性。方差分析结果显示,47项表型指标中,基部第3节和穗位节在不同自交系中差异最显著的指标均为周皮区类表型指标,进一步表明基于区带的维管束解析方法相较传统手段更有助于玉米茎秆维管束表型的精准鉴定。表型指标从基部到顶部的分布趋势研究表明,大部分表型指标分布趋势一致,均呈依次减小的趋势,但功能区(表皮区/周皮区/髓区)面积占比却不一致,表明基于区带的维管束解析方法获取的表型指标更能突显出不同节间差异,进而为后续遗传学分析和性状定向改良提供表型参考。
参考文献:
[1]赵春江. 植物表型组学大数据及其研究进展[J]. 农业大数据学报,2019,1(2):5-18.
[2]胡伟娟,傅向东,陈 凡,等. 新一代植物表型组学的发展之路[J]. 植物学报,2019,54(5):558-568.4FC70124-2524-41CF-BEF5-849E79D3D36F
[3]张 颖,廖生进,王璟璐,等. 信息技术与智能装备助力智能设计育种[J]. 吉林农业大学学报,202 43(2):119-129.
[4]Huang C,Chen Q Y,Xu G H,et al. Identification and fine mapping of quantitative trait loci for the number of vascular bundle in maize stem[J]. Journal of Integrative Plant Biology,2016,58(1):81-90.
[5]Ren H,Jiang Y,Zhao M,et al. Nitrogen supply regulates vascular bundle structure and matter transport characteristics of spring maize under high plant density[J]. Frontiers in Plant Science,202 11:602739.
[6]馮海娟,张善平,马存金,等. 种植密度对夏玉米茎秆维管束结构及茎流特性的影响[J]. 作物学报,2014,40(8):1435-1442.
[7]杨 硕,郑云霄,黄亚群,等. 不同玉米自交系茎秆上部维管束数目的差异分析[J]. 植物遗传资源学报,202 2(2):466-475.
[8]Legland D,Devaux M F,Guillon F. Statistical mapping of maize bundle intensity at the stem scale using spatial normalisation of replicated images[J]. PLoS One,2014,9(3):e90673.
[9]Heckwolf S,Heckwolf M,Kaeppler S M,et al. Image analysis of anatomical traits in stalk transections of maize and other grasses[J]. Plant Methods,2015,11(1):1-11.
[10]Legland D,El-Hage F,Méchin V,et al. Histological quantification of maize stem sections from FASGA-stained images[J]. Plant Methods,2017,13(1):1-11.
[11]赵 欢,王璟璐,廖生进,等. 基于Micro-CT的玉米籽粒显微表型特征研究[J]. 智慧农业(中英文),202 3(1):16-28.
[12]Shao M,Zhang Y,Du J J,et al. Fast analysis of maize kernel plumpness characteristics through Micro-CT technology[C]// Rannenberg K. International Conference on Intelligent Agriculture 017(ICIA2017) PartI. Changchun: Beijing Engineering Research Center for Agriculture Internet of Things, China Agricultural University,2017:46-54.
[13]Zhang Y,Ma L M,Pan X D,et al. Micron-scale phenotyping techniques of maize vascular bundles based on X-ray microcomputed tomography[J]. Journal of Visualized Experiments,2018 (140):e58501.
[14]Du J J,Zhang Y,Guo X,et al. Micron-scale phenotyping quantification and three-dimensional microstructure reconstruction of vascular bundles within maize stalks based on micro-CT scanning[J]. Functional Plant Biology,2016,44(1):10-22.
[15]Wu D,Wu D,Feng H,et al. A deep learning-integrated micro-CT image analysis pipeline for quantifying rice lodging resistance-related traits[J]. Plant Communications,202 (2):100165.
[16]Zhang Y,Ma L M,Wang J,et al. Phenotyping analysis of maize stem using micro-computed tomography at the elongation and tasseling stages[J]. Plant Methods,2020,16(1):1-14.
[17]Zhang Y,Du J J,Wang J L,et al. High-throughput micro-phenotyping measurements applied to assess stalk lodging in maize (Zea mays L.)[J]. BioMed Central,2018,51(1):40.4FC70124-2524-41CF-BEF5-849E79D3D36F
[18]Zhang Y,Wang J L,Du J J,et al. Dissecting the phenotypic components and genetic architecture of maize stem vascular bundles using high‐throughput phenotypic analysis[J]. Plant Biotechnology Journal,202 19(1):35-50.
[19]Yang X H,Gao S B,Xu S T,et al. Characterization of a global germplasm collection and its potential utilization for analysis of complex quantitative traits in maize[J]. Molecular Breeding,201 8(4):511-526.
[20]何启平,董树亭,高荣岐. 不同类型玉米品种果穗维管束的比较研究[J]. 作物学报,2007,33(7):1187-1196.
[21]李素坤,张秋芝,郝玉兰,等. 玉米成熟期茎秆石蜡切片方法的研究[J]. 安徽农业科学,2010,38(8):3935-3937.
[22]Zhang Y,Guo X Y,Yan W P,et al. Three-dimensional visualization of vascular bundles in stem nodes of maize[J]. Fresenius Environmental Bulletin,2017,26(5):3395-3401.
[23]張士龙,贾海涛,贺正华,等. 长江中游夏玉米成熟期茎秆力学特性及其影响因素研究[J]. 现代农业科技,2020(24):9-12,17.
[24]蒋傲男,闫静琦,卢海博,等. 不同春玉米品种茎秆显微结构对抗折强度的响应[J]. 玉米科学,2020,28(5):53-59.
[25]李 波,张吉旺,崔海岩,等. 施钾量对高产夏玉米抗倒伏能力的影响[J]. 作物学报,2012,38(11):2093-2099.
[26]何启平,董树亭,高荣岐. 玉米果穗维管束系统的发育及其与穗粒库容的关系[J]. 作物学报,2005,31(8):995-1000,1105-1106.
[27]侯彦龙,马 丹. 玉米生长发育规律研究现状[J]. 中国农业信息,2014(23):14.
[28]王 越,李小雨,李青青,等. 不同杂种优势群玉米茎秆维管束性状比较分析[J]. 河北农业大学学报,202 44(5):14-20,56.
[29]赵 雪,周顺利. 玉米抗茎倒伏能力相关性状与评价研究进展[J]. 作物学报,2022,48(1):15-26.4FC70124-2524-41CF-BEF5-849E79D3D36F
猜你喜欢
石油实验地质(2022年6期)2022-12-07 02:14:48
天然气技术与经济(2022年1期)2022-03-21 05:48:14
河北北方学院学报(自然科学版)(2022年11期)2022-02-03 06:04:56
医疗装备(2020年13期)2020-08-24 07:30:06
国际检验医学杂志(2019年17期)2019-09-10 02:44:26
石油工业技术监督(2019年7期)2019-07-30 07:59:04
特种油气藏(2018年6期)2019-01-11 02:51:16
石油与天然气地质(2015年2期)2015-05-15 03:34:35
长江大学学报(自科版)(2014年2期)2014-03-20 13:20:26
中国石油大学学报(自然科学版)(2014年2期)2014-02-28 16:12:27
