
利用生物信息学探讨IL-8在慢性阻塞性肺疾病中异常表达及其相关基因的功能
2022-06-22 08:05:32张平安高娜陈明哲李潇宁纪国超吴建军
广东药科大学学报 2022年3期
张平安,高娜,陈明哲,李潇宁,纪国超,吴建军*
(1.北京中医药大学第三附属医院,北京 100029;2.河南中医药大学,河南 郑州 450046)
慢性阻塞性肺疾病(chronic obstructive pulmo‐nary disease,COPD)简称慢阻肺,是以持续呼吸道症状和气流受限为特征的慢性气道炎症疾病,发病过程中外周气道、肺实质和肺血管中巨噬细胞、中性粒细胞以及包括Tc1、Th1、Th17和ILC3淋巴细胞等炎性细胞明显增加,并释放多种炎症介质[1]。COPD 因其逐年升高的发病率,正逐渐成为影响人们健康的慢性疾病。2017 年全球疾病负担研究指出,全球慢阻肺患者数约3 亿人,从2007 年到2017年增长了24.9%[2]。研究证明COPD 存在持续的气道与全身炎症反应,炎性介质、免疫细胞以及趋化因子的水平可以预测COPD 的急性加重及预后[3]。而白介素8(IL-8)和肿瘤坏死因子受体(TNF-α),在AECOPD(acute exacerbation of COPD)和COPD 患者血清中的表达升高并且与COPD的严重程度呈正相关[4]。通过分析COPD 患者连续3 年每年发生≥1次AECOPD 的个体相关因素发现,COPD 频繁急性加重表型与CT 表现的小气道异常和白细胞介素8(IL-8)有关[5]。由此可知,IL-8可能通过参与炎症反应而诱导慢阻肺的急性加重,而调控IL-8 则可能成为治疗COPD的潜在或有效途径之一。
2011年,有研究提出竞争性内源RNA(ceRNA)可以通过其miRNA反应元件作为miRNA海绵以调节其表达的假设[6]。近年有研究证明lncRNAs 具有ceRNAs 作用,并且在恶性和慢性疾病中发挥重要作用[7]。经研究发现miR-122-5p-A2M-LINC00987相互作用可能在COPD 中发挥重要作用[8],而miR-122-5p 又可能通过调节人血浆蛋白酶抑制剂(A2Mα2-巨球蛋白)以调节肺蛋白酶活性[9]。研究发现作为miRNA 调控的下游基因,白细胞介素(IL-8)可以通过调控ROCH 炎症通路诱导气道高反应性和促进气道平滑肌痉挛,加重肺功能损害[10]。
但是目前IL-8在COPD的发病中其调控机制仍旧不明,因此本文拟通过GEO数据库筛选与IL-8表达相关的转录组数据,探究IL-8表达调控相关的作用机制。
1 材料与方法
1.1 数据来源与预处理
本研究所采用的基因表达数据来源于GEO 基因表达综合数据库(https://www.ncbi.nlm.nih.gov/geo/)。慢阻肺患者和正常对照组mRNA 数据来源于GSE5058、GSE38974,miRNA 数 据 来 源 于GSE24709,lncRNA数据来源于GSE107426。
1.2 独立秩和检验比较IL-8的表达差异
利 用R3.5.2 软 件 的limma 包 对GSE5058、GSE38974 数据的IL-8 表达量进行独立样本秩和检验,而Wilcoxon 秩和检验是一种流行的非参数检验,用于比较2 个独立的总体的差异以筛选差异基因,数据格式为慢阻肺患者和正常对照组模式[11-12]。
1.3 基因富集分析
将GSE5058、GSE38974 数据中的慢阻肺患者按照IL-8 的表达量进行从高到低排序。按照IL-8表达量的中位数将患者分IL-8 高表达组和低表达组。本研究用GSEA(version 3.0)软件对表达矩阵进行分析,采用c5.all.v7.2.symbols.gmt 数据集,按照缺省参数设置进行GSEA,设定随机组合次数为1000,|NES|>1,P<0.05 的基因集被认为是显著富集的。
1.4 筛选IL-8 高表达的正负相关的差异基因,筛选差异miRNA、lncRNA,绘制IL-8环状网络图
利用微生信在线数据平台(http://www.bioin‐formatics.com.cn)绘制upset 图,对GSEA 基因富集分析的GSE5058、GSE38974 数据中与IL-8 高表达呈正、负相关的基因取交集;利用R3.5.2软件的相关limma 包对miRNA 数据集GSE24709 数据进行差异miRNA 的分析,利用lncRNA 数据集GSE107426 筛选差异lncRNA,筛选条件为log2foldchange<-1,P<0.05 的miRNA 被认为是显著低表达;并利用miRWalk(http://mirwalk.umm.uni⁃heidelberg.de/)筛选与IL-8 相互作用的miRNA;利用StarBase3 平台(http://starbase.sysu.edu.cn/)筛选与miRNA 互相作用的lncRNA,然后利用Cytoscape3.7.1 软件绘制IL-8-miRNA-lncRNA网络图。
1.5 利用String 平台筛选IL-8 高表达呈正负相关的关键基因
将IL-8 高表达呈正、负相关的基因输入String平台(https://string-db.org/)构建蛋白互作网络(pro‐tein-protein interaction,PPI),选取“Homo sapiens”为蛋白属种,最低相互作用阈值“medium confidence”(>0.15),其他参数设置为默认参数,获取PPI蛋白互作网络,利用Cytoscape3.7.1 软件中MOCDE 插件进行关键基因的筛选,筛选条件为Node Score Cutff:0.2,K-Core:2,Max Depth:100。
1.6 关键基因进行GO和KEGG富集分析
利用R3.5.2软件中的clusterProfiler包对筛选出的IL-8 高表达呈正、负相关的关键基因进行GO 和KEGG富集分析,筛选条件为矫正后的P值<0.05。
2 结果
2.1 GEO样本数据统计
通过GEO 数据库筛选出符合慢阻肺患者和正常对照组对比的mRNA 数据GSE5058 数据包含慢阻肺患者15 例,正常对照组患者24 例,GSE38974数据包含慢阻肺患者23 例,正常对照组患者9 例;miRNA 数据集GSE49881 数据包含慢阻肺患者41 例,正 常 对 照 组 患 者16 例;lncRNA 数 据 集GSE107426 包含慢阻肺患者1 例,正常对照组患者1例。如表1所示。
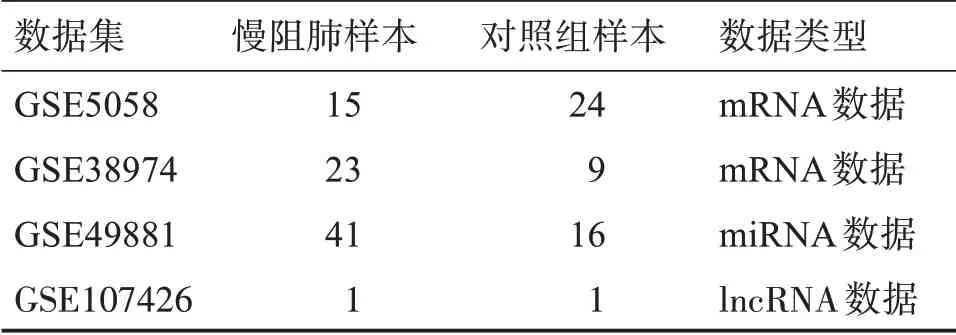
表1 数据集及分布情况Table 1 Data set and distribution
2.2 慢阻肺患者和正常对照组IL-8表达差异
通过对慢阻肺患者和正常对照组IL-8 表达的水平发现,IL-8 在慢阻肺患者中显著高表达。其中GSE5058 数据中IL-8 的P值为0.000331,log2fold‐change=1.3084674,如图1A 所示;GSE38974 数据 中IL-8 的P值 为0.000418,log2foldchange=2.2547985,如图1B所示。
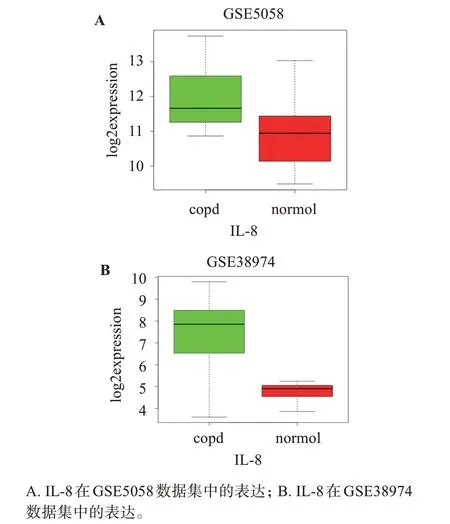
图1 IL-8在GEO数据集中的表达Figure 1 Expression of IL-8 in GEO dataset
2.3 GSEA基因富集分析结果
利用GSE5058 和GSE38974 这2 个数据集中的慢阻肺患者基因表达矩阵,分别计算IL-8 与其他基因表达的相关性。GSEA 分析发现有47 条通路显著富集在IL-8 高表达患者中。如表2 所示,高IL-8患者富集的通路主要涉及细胞转化和一些受体信号通路。对排名为前2 的通路进行了展示(图2 A、B)。其中GSE5058 数据集 筛选到314 个显著与IL-8 表达相关的基因(147 个正相关,167 个负相关)。GSE38974 数据集筛选到2239 个显著与IL-8 表达相关的基因(692个正相关,1547个负相关)。

图2 GSEA基因富集分析IL-8高表达显著相关通路top2展示Figure 2 Top 2 pathways related to high expression of IL-8 by GSEA gene enrichment analysis
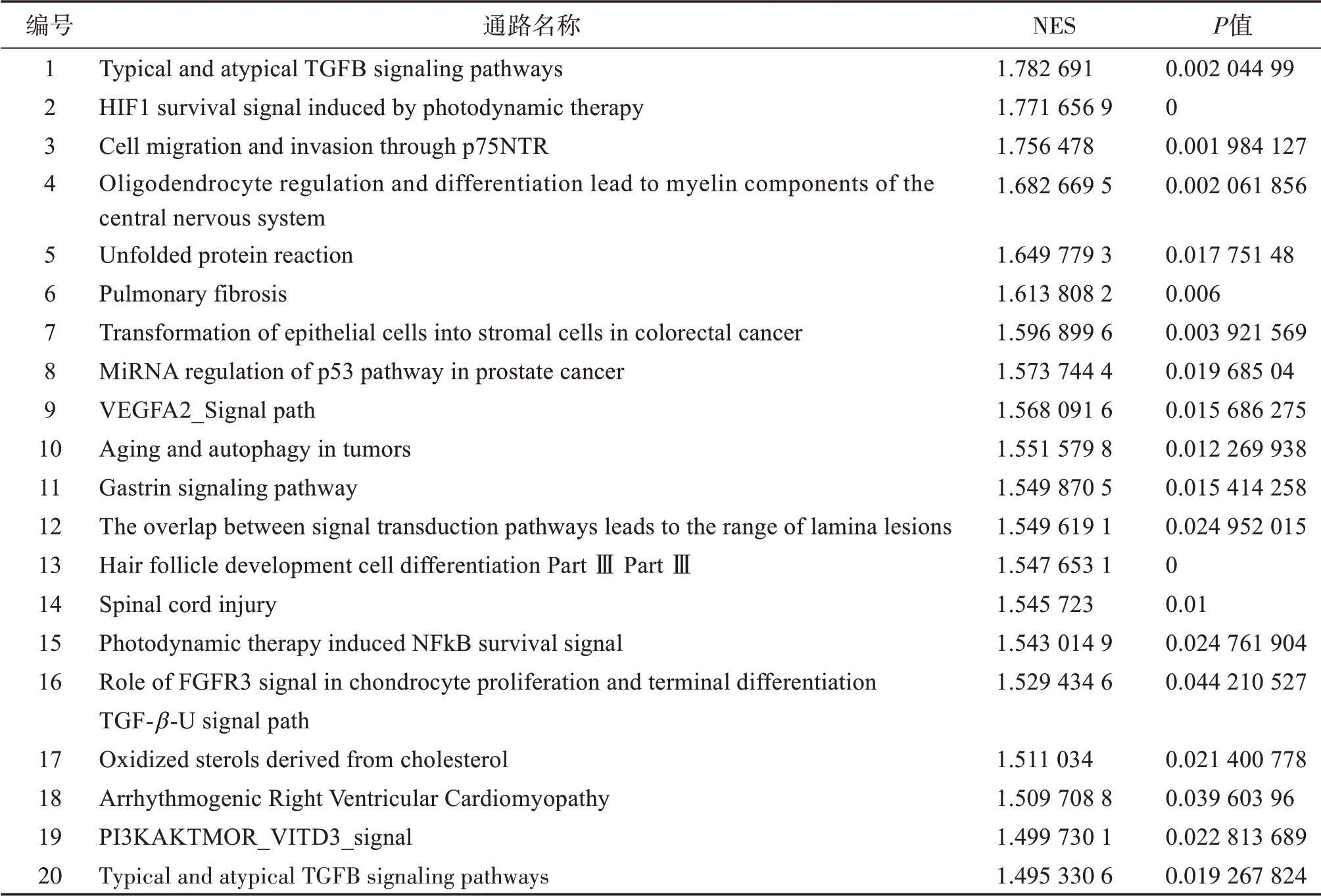
表2 IL-8高表达相关的前20条通路Table 2 Top 20 pathways related to high expression of IL-8
2.4 筛选IL-8 高表达呈正、负相关的基因,筛选差异miRNA,绘制相关网络图
通过微生信在线数据平台(http://www.bioin‐formatics.com.cn)绘制upset 图,对GSEA 基因富集分析的GSE5058、GSE38974 数据集中IL-8 高表达呈正、负相关的基因取交集,一共得到111 个相关基因(36 个正相关,75 个负相关,如图3 A、B 所示);利用筛选得到的与IL-8相关的miRNA和miRNA相关的lncRNA 在Cytoscape3.7.1 软件中绘制网状图,如图3 C、表3所示,其中miRNA与mRNA呈负调控的关系;mRNA与lncRNA可竞争性作用于miRNA。

表3 差异miRNA和差异lncRNA表达结果Table 3 Differential miRNA and lncRNA expression results
2.5 利用String 平台筛选差异基因蛋白互作网络中的关键基因
通过String 平台将IL-8 高表达呈正、负相关的基因的PPI 蛋白互作网络并根据筛选条件得到IL-8表达相关的正相关基因有PTGS2、STAT3、IL1B、TNF、VEGFA等20 个;与IL-8 表达相关的负相关基因有PRKCZ、JUN、TGFB1、TP53、MAPK1等17 个,如图3D、E所示
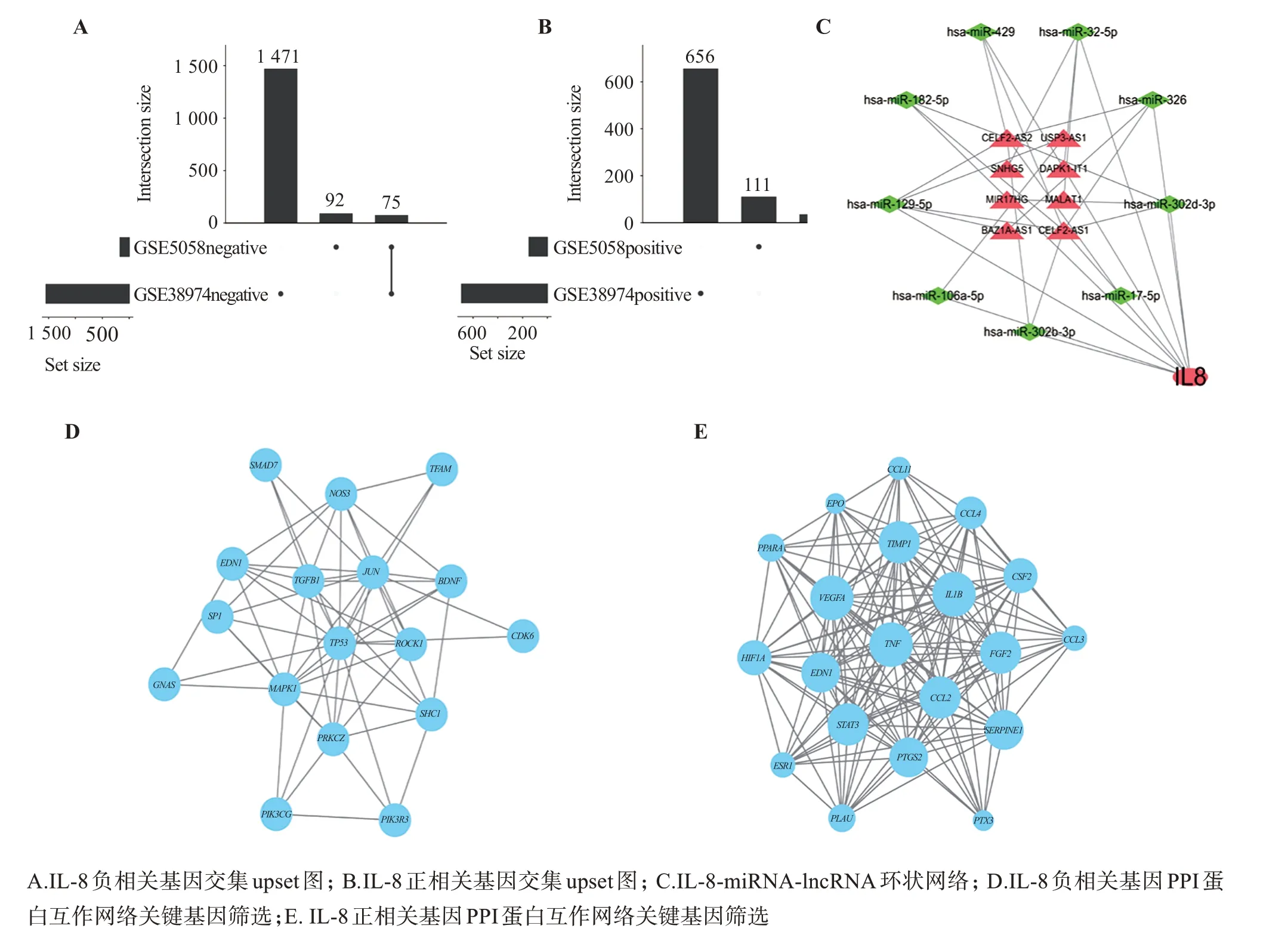
图3 IL-8正负相关基因、IL-8-miRNA-lncRNA环状网络、IL-8正负相关基因PPI网络关键基因筛选Figure 3 IL-8 positive and negative related genes,IL-8-miRNA-lncRNA ring network,and screening of key genes in PPI network of IL-8 positive and negative related genes
2.6 差异基因进行GO和KEGG富集分析
选取IL-8 表达正相关的基因进行GO 和KEGG富集分析,以分析IL-8 表达促进的功能(如图4 C 所示)或通路(如图4 D所示)。在GO功能方面上主要涉及细胞因子活性(cytokine activity)、G 蛋白偶联受体结合(G protein-coupled receptor binding)、激素活性(hormone activity)、激素受体结合(hormone receptor binding)、趋化因子受体结合(chemokine receptor binding)等通路的调控;KEGG 富集分析集中在IL-17信号通路(IL-17 signaling pathway)、肿瘤坏死因子信号通路(TNF signaling pathway)、NF-κB信号通路(NF-kappa B signaling pathway)、细胞因子-细胞因子受体相互作用(cytokine-cytokine recep‐tor interaction)等通路上。选取IL-8 表达负相关的基因进行GO 和Kegg 富集分析,以分析IL-8 表达抑制的功能(如图4 A所示)或通路(如图4B所示)。在GO功能方面上主要涉及细胞因子受体结合(cyto‐kine receptor binding)、转化生长因子β受体结合(transforming growth factor beta receptor binding)、组蛋白乙酰转移酶结合(histone acetyltransferase binding)、组蛋白脱乙酰酶结合(histone deacetylase binding)、磷脂酰肌醇3-激酶活性(phosphatidylinosi‐tol 3-kinase activity)等功能;KEGG 富集分析集中在非小细胞肺癌(non-small cell lung cancer)、HIF-1信号通路(HIF-1 signaling pathway)、PI3K-Akt 信号通路(PI3K-Akt signaling pathway)等通路中。
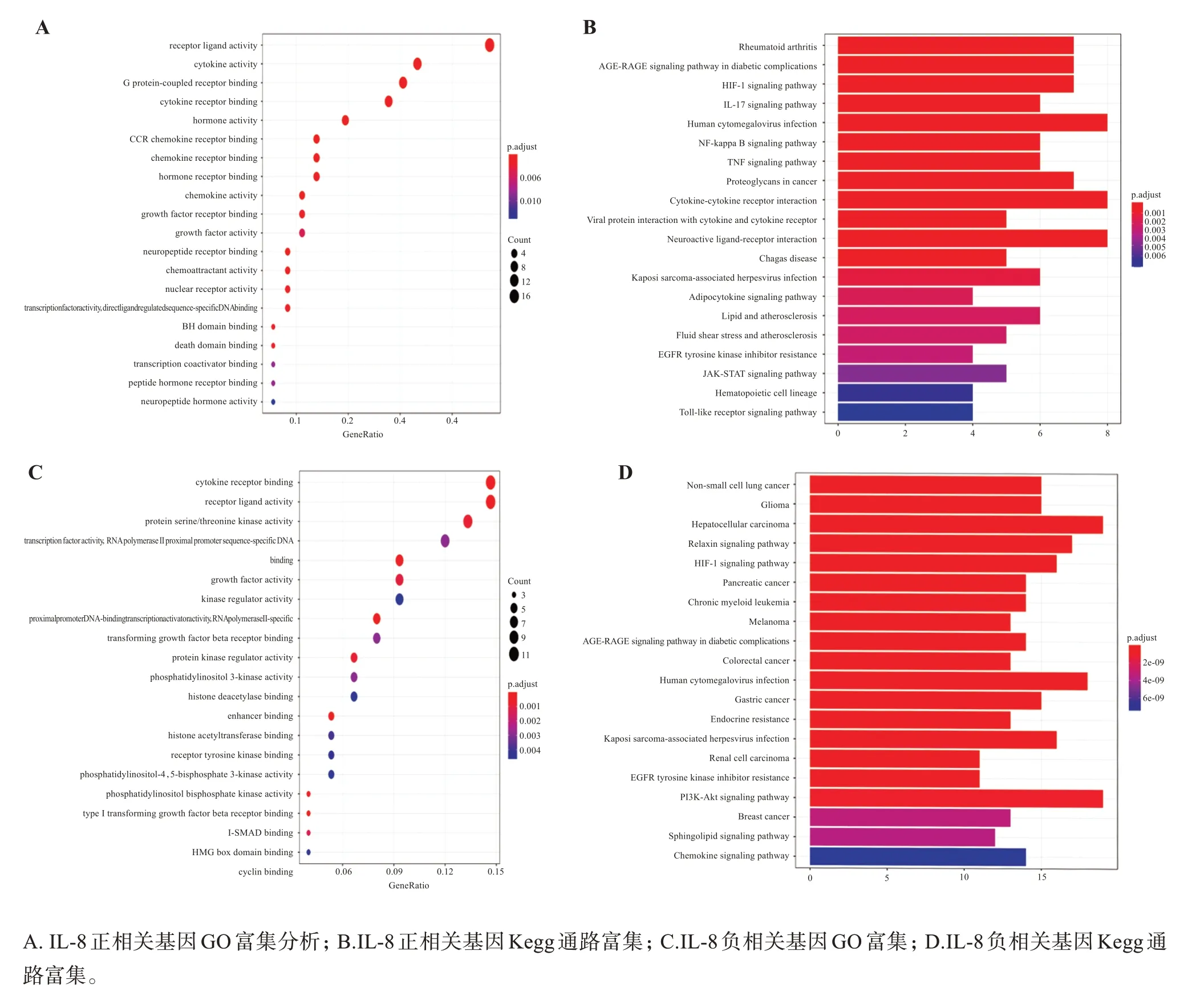
图4 IL-8正负相关基因GO富集和Kegg通路富集结果Figure 4 GO enrichment and KEGG pathway enrichment of IL-8 positive and negative related genes
3 讨论
IL-8 是一种有效的中性粒细胞募集和激活因子,与许多炎性肺病(急性呼吸窘迫综合征、慢性阻塞性肺疾病、哮喘)的发病机制有关,其机制通过释放弹性蛋白酶而损伤肺功能[13],且有研究通过酶联免疫吸附试验发现COPD 患者获得的痰液标本中IL-8 水平升高[14]。而通过对比COPD 和正常人之间的转录组表达数据发现,IL-8 在COPD 患者中显著高表达,说明IL-8的高表达与COPD发病相关。
目前许多研究已经证明lncRNA 可以与mRNA竞争性结合miRNA,以调控mRNA 的表达[15]。本研究发现环状通路中hsa-miR-106a-5p、hsa-miR429、hsa-miR-32-5p 等可以调控IL-8 的表达,而这一过程可 以 被USPS-AS1、CELF2-AS1、CELF2-AS2、SN‐HG5、DAPK1-1T1、MALAT1等lncRNA 竞争性抑制。研究表明miR-32-5p 可能通过上调NFIL3 及抑制炎性因子TNF-α、IL-6 的表达以抑制脂多糖诱导的肺上皮细胞凋亡和炎性反应[16]。在MicroRNAs 作为COPD 分级标志物的筛选、验证及功能分析中通过实时荧光PCR 方法检测COPD 患者和正常人外周血中miR-106a-5p 的表达,发现miR-106a-5p 在COPD 患者中显著高表达,在COPD 的发生发展中起重要作用,与COPD 的严重程度相关[17]。有研究证明lncRNA SNHG5 可以通过SNHG5-miR-132-PTFN 轴调控miR-132 下游靶标PTEN 蛋白的表达,以减轻香烟烟雾刺激对COPD人支气管上皮细胞的增殖、凋亡和炎症(IL-1β、IL-6 和TNFα)反应[18]。还有研究表明lncRNA-MALAT1 可以通过调节MALAT1/miR-146a/COX2 轴,延缓COPD 肺功能的减低,并且可以减轻COPD 的慢性炎症,有望成为预测COPD 严重程度的新型生物标志物[19]。结合IL-8-miRNA-lncRNA 环状网络可以得知,lncRNA中SNHG5、MALAT1可以通过竞争性结合miR-32-5p,发挥内源性竞争IL-8 以调节IL-8 在COPD 中的作用,因此推测lncRNA 中SNHG5、MALAT1 可以通过SNHG5/MALAT1-miR-32-5p-IL-8 轴调控IL-8在COPD中的炎症过程。
通过PPI蛋白互作网络分析得知,与IL-8高表达呈正相关的基因为TNF、VEGFA、PTGS2、STAT3、IL1B等蛋白,与IL-8高表达呈负相关的基因为PRK‐CZ、JUN、TGFB1、TP53、MAPK1等蛋白;其中肿瘤坏死因子(TNF-α)作为COPD中的促黏附细胞因子,在COPD患者的血浆和痰液中明显升高[20],并且经研究发现TNF-α多态性基因表型可以影响慢性阻塞性肺疾病的临床表型(增加发病率,使EV1下降增快、体重指数降低),此过程与肺分泌过表达TNF-α有关,并伴有IL-8等炎症因子的过表达和增加的中性粒细胞迁移[21]。研究表明血管内皮生长因子(VEGFA)在COPD 中显著高表达并参与人肺微血管细胞的凋亡过程,miRNA-206可以诱导VEGFA的过表达,这一过程被miRNA-206 敲低显著逆转[22]。TP53 是一种应激反应基因,通过激活下游基因的转录发挥基因毒性应激、致癌信号、细胞损伤。研究发现TP53 持续下调及其下游CDKNIA通路可能是通过COPD向鳞状细胞癌(SQCC)发展,与抑制细胞周期停滞和细胞凋亡有关[23],因此推测高表达的IL-8 可能通过调节与其正负相关基因的表达来调控COPD 疾病的发生。
通过IL-8 表达正相关的基因进行GO 和Kegg富集分析发现,调控正相关基因表达的通路也有如下功能:如GC 发挥作用需要与GC 受体结合,形成GC-GR 复合物,而炎性通路的激活则会降低受体的表达,减轻受体的活性,导致糖皮质激素的不敏感[24],而通过对慢阻肺疾病激素不敏感与糖皮质激素受体核内移的相关性研究中发现,COPD 患者外周血中GRα的核内转移与GC 的敏感性呈正相关,而糖皮质激素的减弱可能与GRα的核内转移受抑制/核内GRα的降解有关[25]。还有研究表明,TNF-α/IL-17 信号通路可以靶向调控IL1B的表达,进而促进CSE 刺激引起的慢阻肺支气管上皮细胞的增殖,并抑制其凋亡以干预COPD 的发展[26]。此外通过研究发现核因子(NF)κB 可以调节细胞因子、趋化因子、生长因子、细胞黏附因子等多种靶基因的表达,进而调控各种组织细胞的炎症与免疫反应、细胞凋亡及增殖分化等生理过程[27],而NF-κB 信号通路的异常激活可以诱导TNF-α、IL-1β、IL-6 以及黏附分子等炎症介质的释放,从而加重COPD的发生,通过抑制其通路可以阻断COPD的进展[28]。
通过IL-8 表达负相关的基因进行GO 和Kegg富集分析发现,调控负相关基因表达的通路有如下功能:慢性阻塞性肺疾病中持续存在的氧化应激反应可以降低肺组蛋白乙酰酶活性,打破组蛋白乙酰转移酶/组蛋白脱乙酰酶的平衡,进而导致组蛋白分子过乙酰化、增强炎症蛋白基因的转录和炎症蛋白的合成,最终能够阻断糖皮质激素的抗感染效应[29]。另有研究发现,PI3K-Akt 通路可干预HDAC2 的表达或活性,其机制为氧化应激反应激活PI3K/Akt 通路,使得该通路下游的信号蛋白Akt发生磷酸化,进一步影响其下游的HDAC2,引起HDAC2 活性下降或失活,导致炎症基因的转录和炎症因子的合成加强[30-32]。还有研究发现小青龙汤中槲皮素等成分可以通过HIF-1 信号通路调控IL-6 等炎症因子的释放以干预COPD 发生发展中的氧化应激、炎症反应过程[33]。结合IL-8 表达正负相关基因的GO 和KEGG的研究发现,与IL-8 表达正负相关的基因可能通过调控组蛋白乙酰转移酶结合、组蛋白脱乙酰酶结合、激素活性和激素受体等生物过程,以及PI3KAkt 通路、HIF-1 信号通路、TNF-α/IL-17 信号通路、NF-kB 信号通路等信号通路在COPD 的发生过程中抑制糖皮质激素敏感性、参与炎症反应。
本研究利用生物信息学的研究方法,发现IL-8在COPD 疾病中显著高表达,并且通过筛选miRNA和lncRNA 得到了IL-8-miRNA-lncRNA 环状网络,采用GSEA 软件筛选出与高表达相关的正负相关mRNA,采用R 软件发现部分GO 生物学过程和KEGG 通路涉及糖皮质激素抵抗和炎症反应的机制。本研究显示,IL-8 有望成为治疗COPD 的新靶点。
猜你喜欢
天津医科大学学报(2021年1期)2021-12-05 11:11:05
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
中国医学影像技术(2017年11期)2017-01-16 12:54:05
哈尔滨医药(2016年3期)2016-12-01 03:58:33
考试周刊(2016年63期)2016-08-15 14:33:26
作文教学研究(2016年1期)2016-07-05 12:22:47
科技视界(2016年1期)2016-03-30 14:08:41
癌变·畸变·突变(2016年3期)2016-02-27 06:15:25
医学研究杂志(2015年8期)2015-06-22 14:00:57
