
基于叶绿体基因组的木兰科植物系统发育探究
2022-06-20 02:09王晓双刘映良
生物学杂志 2022年3期
朱 斌,钱 方,王晓双,刘映良
(贵州师范大学 生命科学学院,贵阳 550025)
木兰科(Magnoliaceae)是双子叶植物中较为原始的科,基于不同的分类系统,木兰科有广义和狭义之分[1]。在恩格勒(Engler)系统分类中,广义的木兰科包括木兰亚科、八角亚科、五味子亚科、水青树亚科等[2]。在哈钦松(Hutchinson)系统中,狭义的木兰科指木兰亚科和鹅掌楸亚科[3]。狭义的木兰科下有18属,约335种,我国有14属,约165种,主要分布于我国东南部至西南部[4]。木兰科植物不仅是温带森林的重要组成成分[5],也是重要的观赏和药用植物[6],木兰科植物具有较为原始的性状,常被作为探究被子植物起源与进化的典型代表物种[7-8]。
叶绿体(chloroplast, cp)是绿色植物进行光合作用的场所,普遍认为起源于“内共生”的古蓝细菌[9]。叶绿体拥有独立的遗传物质[10]。由于叶绿体基因组具有相对保守、进化速率适中、基因组小等特点[11-12],因此cpDNA被广泛应用于解决物种间的系统进化关系[13-14]。此外叶绿体蛋白编码序列也可用于植物的系统发育分析[15]。matK基因是位于叶绿体赖氨酸trnA基因(trnK)的内含子内、单一拷贝编码基因,是叶绿体基因组中蛋白编码基因中进化速率最快的基因之一[16],对系统学的研究具有重要价值。采用单个基因序列分析物种间的系统关系已应用于许多科属中,如鳞毛蕨科[17]、石斛属[18]和木犀属[19]等。
不同的分类学者从形态学、解剖学和孢粉学等对木兰科进行了系统划分,但除鹅掌楸属外,分类学者对木兰科其他属的划分存在很大分歧[20]。如Nooteboom[21]认为华盖木属、拟单性木兰属应归并为木兰属,而合果木属和观光木属应并入含笑属;随后通过对中国木兰科植物的分类研究,Chen 等[22]又将华盖木属归并入木莲属;有学者通过对木兰亚科主要性状的观察,将中国木兰亚科分为木兰属和含笑属,而合果木属、玉兰亚属及观光木属被并入含笑属;王猛等[23]通过ISSR分子标记技术对分布于云南的木兰科植物进行聚类分析,聚类结果发现传统的分类系统存在差异,含笑属、木兰属和木莲属间存在明显属间交叉现象。总体而言,木兰科植物的分类分歧主要存在于木兰亚科下的一些单种属和寡种属之间。由于测序技术的限制,前人只是对极少数的木兰科物种基因序列进行系统发育分析,鲜有cpDNA水平的系统发育研究。本研究基于已公布的广义木兰科41个物种进行系统发育分析,利用叶绿体全基因组序列(WCGS)、编码序列(CDS)和matK序列构建系统发育树,为木兰科系统发生关系的构建提供科学依据,以及为该科物种精准鉴定和系统发育研究提供参考,为木兰科植物的种质资源保护与开发提供科学依据。
1 材料和方法
1.1 材料
从NCBI(https://www.ncbi.nlm.nih.gov/)数据库共获取了41个已公布的木兰科物种的叶绿体基因组(数据获取的截止日期为2020年11月15日),以拟南芥(Arabidopsisthalian)叶绿体基因组作为外类群,木兰科物种叶绿体基因组信息和GenBank登录号见表1。
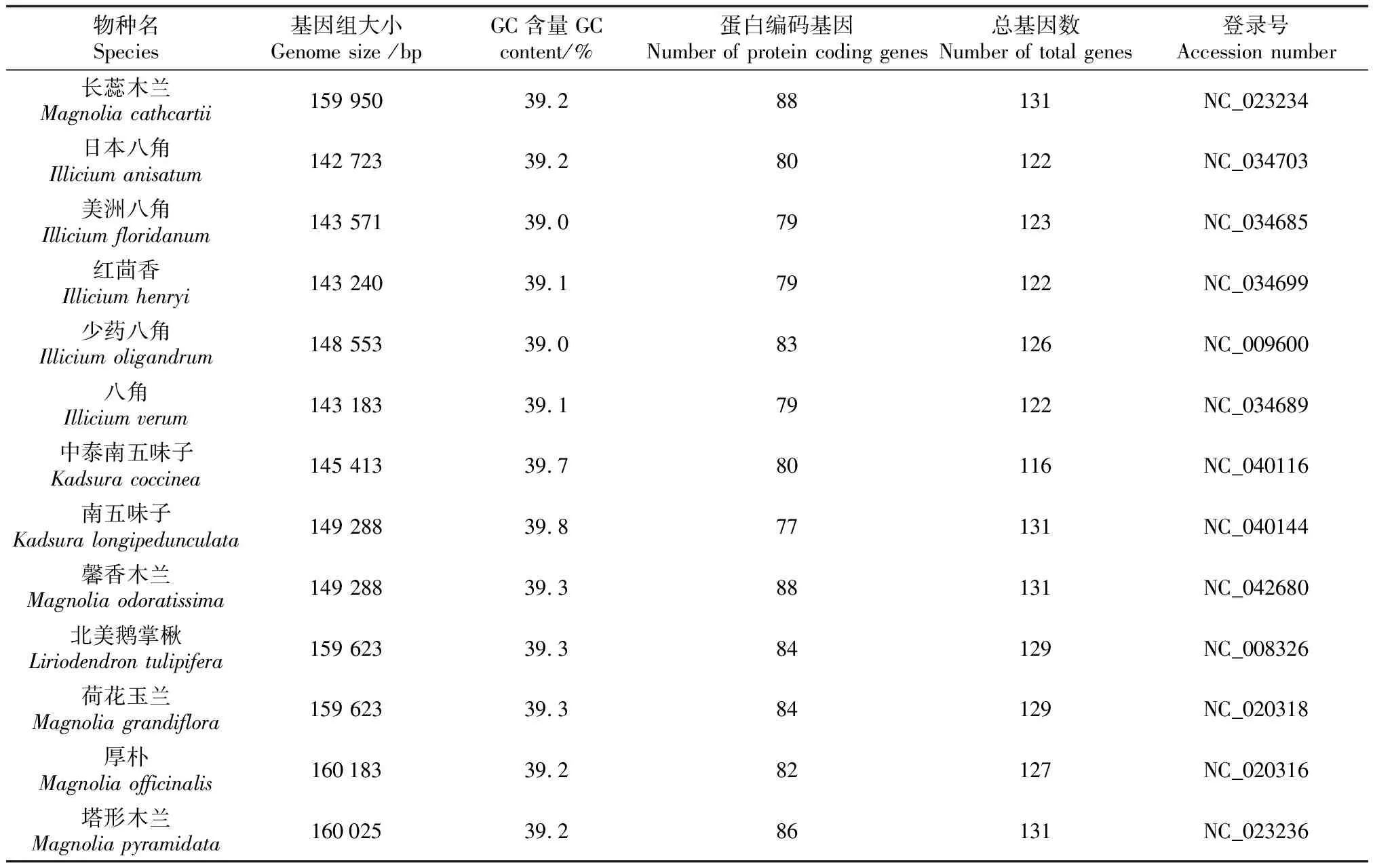
表1 研究所用的木兰科植物及拟南芥叶绿体基因组信息
1.2 系统发育树构建
将数据经EditPlus进行初步整理,直接下载WCGS和matK基因序列处理之后用MEGA 7.0以Maximum Likelihood(ML)法构建系统发育树;将从NCBI下载经过初步筛选处理的注释文件,选择所有物种的蛋白编码基因名,通过在线网站http://bioinformatics.psb.ugent.be/webtools/Venn/提取41个木兰科物种的56个共有蛋白编码基因,分别为psbB、ycf3、petG、ycf4、rpoB、psbA、psaJ、ccsA、rps8、rbcL、psbM、rps19、rpl2、psaC、matK、psbE、petA、rpl33、psaA、rpl36、ndhJ、psbK、atpI、psbF、rps2、rpl32、atpH、psbN、psaB、atpA、rps14、infA、psaI、psbJ、rpl16、rpl20、rpoA、ndhD、psbH、rpoC2、ndhA、ndhI、ndhH、atpB、petD、ndhB、atpF、rps7、rpl23、clpP、cemA、ndhE、petL、psbT、rps12和ndhC,使用MEGA7.0以ML法构建系统发育树。
2 结果与分析
2.1 叶绿体基因组基本特征
通过对所选物种的cpDNA比较分析,发现41个木兰科物种的叶绿体基因组大小在142 723~160 183 bp(表1),最小的是日本八角,最大的是厚朴;不同的物种之间序列大小差异在0~460 bp,其中南五味子和馨香木兰、墨西哥巨叶木兰和望春玉兰的序列大小均无差异;41个木兰科cpDNA的GC含量在39.0%~39.8%(平均为39.3%),均高于拟南芥cpDNA的GC含量(36.3%),其中含量最高和最低的是南五味子和美洲八角,分别为39.8%和39.0%;蛋白编码基因数为77~91,南五味子的编码基因个数最少,为77个,云南拟单性木兰的蛋白编码基因个数最多,为91个;叶绿体基因组基因总数为116~138个,其中最少的是五味子,最多的是云南拟单性木兰。
2.2 不同建树方法的比较
对木兰科41个物种的WCGS、CDS和matK基因序列构建系统发育树,统计3种建树方法中各个分支的BVs[图1(a)]。系统发育树共生成39个分支,WCGS系统发育树中BVs值低于90%和位于90%~99%的分别有8个和5个,26个分支的BVs达到100%;而在CDS系统发育树和matK系统发育树中BVs值低于90%的分别有11个和30个,位于90%~99%的分别有10个和5个,CDS系统发育树中有18个节点达到100%,matK系统发育树中仅有4个节点达到100%。总体上,基于WCGS构建的系统发育树分支的BVs高于CDS和matK基因序列构建的系统发育树,其中BVs最低的是基于matK构建的系统发育树。
基于3种系统发育建树结果,狭义木兰科下的31个物种大致均与本属的物种聚为一类,且均聚在木兰亚科这一大分支内,因此将木兰亚科植物合并分析(图2~图4)。3种系统发育树对分类明确的4个属在属间和种间分类时与传统的形态学分类结果一致,差异仅为分支的BVs值。其中鹅掌楸属与木兰亚科植物聚为一个分支,显示较近的亲缘关系;五味子属与南五味子属植物聚为一个亚类,八角属植物自成一类。对比3种建树方法发现对分类明确的10个物种,其9个节点BVs均较高[图1(b)]。其中WCGS系统发育树有8个节点BVs达到100%,仅1个节点为87%(图2);CDS系统发育树中6个节点BVs达到100%,位于90%~100%和低于90%的节点分别有2个和1个;在matK系统发育树中仅有4个节点BVs达到100%,位于90%~100%和低于90%的节点分别有2个和3个。
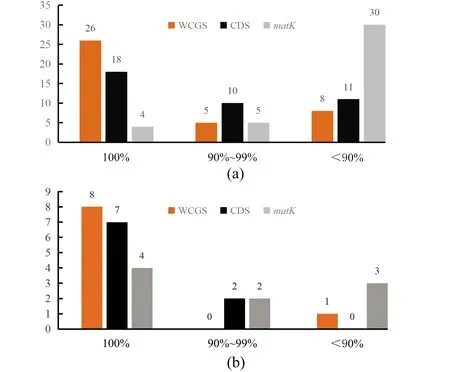
(a)41个物种39个节点的BVs比较;(b)4个明确分类的属的9个节点BVs比较。横坐标代表节点BVs,纵坐标代表节点个数。
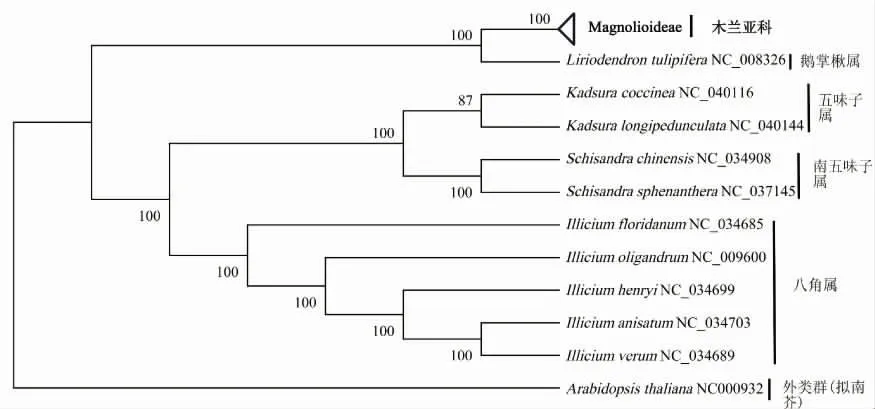
图2 基于木兰科WCGS构建的系统发育树
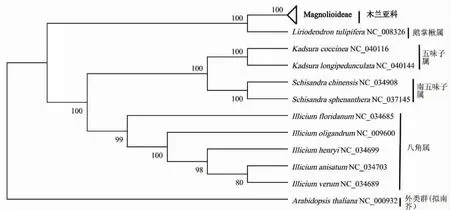
图3 基于木兰科41个物种共有CDS序列构建的系统发育树
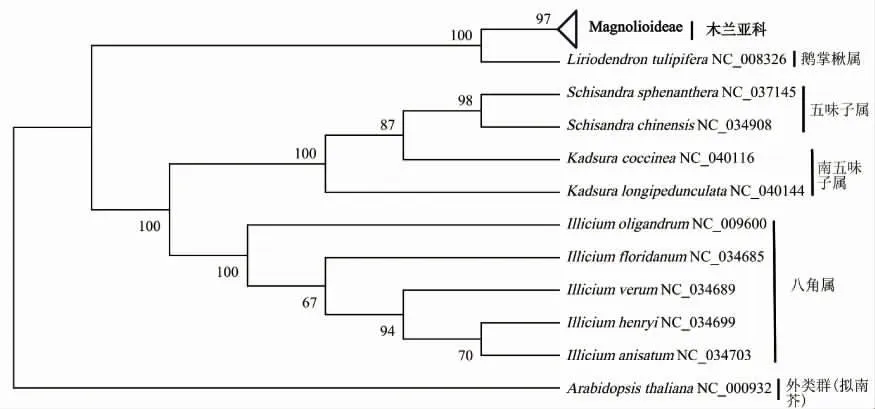
图4 基于matK基因序列构建的系统发育树
2.3 基于WCGS系统发育分析
相较于其他两种方法,WCGS建树法的BVs值更高,为阐释木兰亚科植物间的亲缘关系,对WCGS系统发育树进一步分析(图5)。基于WCGS系统发育树可将31个木兰亚科的物种大致分为7个分支,分别是玉兰属、华盖木属/拟单性木兰属、含笑属、焕镛木属、木兰属、木莲属和喙木兰属。其中喙木兰属(馨香木兰)构成了远离木兰亚科其他物种的一个独立分支。木兰属下的柳叶木兰与玉兰属的物种构成一个分支;华盖木属下的华盖木和拟单性木兰属下的云南拟单性木兰聚成一个独立的分支,与玉兰属构成姐妹群;长蕊木兰和红花木莲聚在含笑属内;焕镛木独立构成一支;木兰属与木莲属组成姐妹类群,荷花玉兰、三瓣木兰、厚朴、西康天女花和天女花聚在木莲属内,其中荷花玉兰为木莲属中较远的一个分支;馨香木兰成为木兰亚科中与其他物种亲缘关系最远的一个独立分支;除南五味子下的两个物种外,其余广义木兰科下的物种均以100%的BVs将各属下物种分别聚在一起。
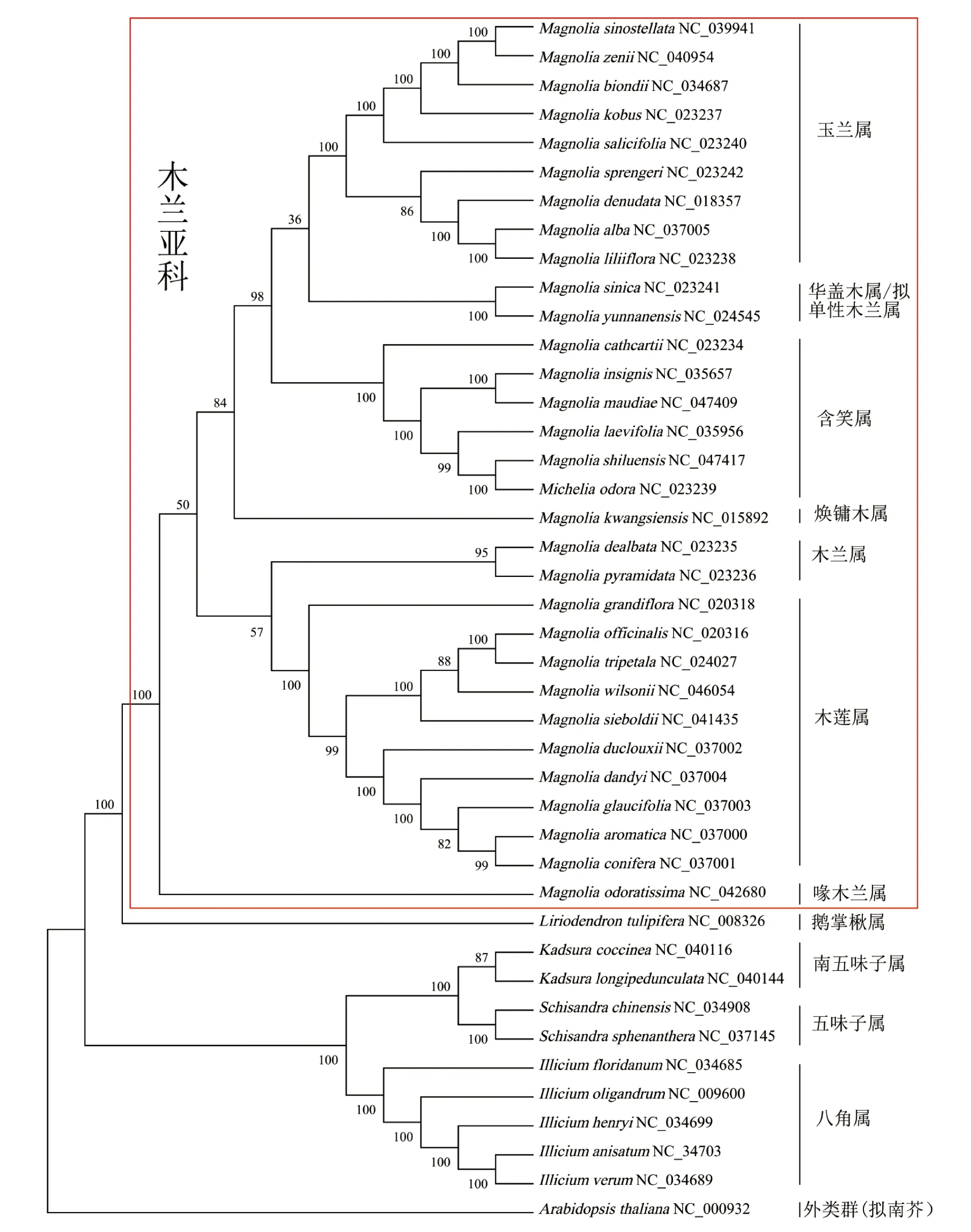
图5 基于WCGS构建的系统发育树
3 讨论与结论
有关木兰科植物的分类研究历经多次整理与修正[24-28],但除鹅掌楸属外,木兰科其他属的划分仍存在很大分歧[20],尤其是对木兰亚科下的一些单种属和寡种属系统分类。究其原因,主要是研究者对木兰科的分类研究多基于形态学、解剖学和孢粉学结果,缺少分子层面的证据支撑。本研究从cpDNA分子层面对木兰科的物种进行了分类探究,系统阐明了41个物种的亲缘关系。木兰科种、属界限的明确将为木兰科濒危植物的保护提供科学依据。同时,掌握木兰科物种的亲缘关系,是木兰科植物种质资源创新、指导引种的关键,有助于木兰科植物的保护与开发。此外,本研究证实了基于叶绿体基因组信息对木兰科植物进行划分的可靠性,可基于叶绿体基因序列开发指纹标签,应用于木兰科植物的资源筛选、鉴定及保护。
此外,3种分类方法均显示木兰亚科与鹅掌楸属亲缘关系相近,五味子科与八角科植物构成两个亲缘关系较近的姊妹类群,支持形态学上对广义木兰科的分类[29]。基于WCGS聚类结果,在木兰亚科中,玉兰属即为木兰属玉兰亚属,柳叶木兰、白玉兰和紫玉兰等被聚在玉兰属之中,赵晶华等[30]通过对atpB和rbcL基因的系统发育分析也证明白玉兰与玉兰属的亲缘关系较近;华盖木为极其濒危的单种属植物[31],被认为是木兰科最原始的植物,传统分类学认为其与木莲属关系较近[32],然而本研究发现华盖木与云南拟单性木兰(云南拟单性木兰属)归为一类,显示二者具有较近的亲缘关系,而与木莲属植物亲缘关系较远,司马永康等[7]系统观察了木兰科植物的幼叶卷叠,也将华盖木属和拟单性木兰属归为一类;焕镛木是我国的特有单属种[33],在系统发育树中单独呈现一个独立的分支,表明了其单属种特性;木莲属与木兰属构成了关系密切的姊妹类群,而木莲属下的两个姊妹分支分别由木兰属和木莲属组成,相对木兰属而言,木莲属是较为原始的类群[34];而红花木莲是木莲属较为原始的物种,与含笑属聚为一类,与传统形态学的分类结果不一致。孟爱平等[35]通过核型分析发现木莲属几个物种的染色体存在一定的差异,推测部分木莲属的植物在进化过程中与含笑属植物经历过胞质基因组的交流,或者非人为干预情况下,生活在同一环境下的不同物种,长期的自然选择可能会出现趋同进化的现象,地域分布比较近则普遍亲缘关系比较近,地域分布较远的物种则可能出现亲缘关系较远的现象。这一现象在研究者对云南分布的木兰科植物的系统进化分析中得到证实[23]。
本研究证实3种聚类方法对分类明确的广义木兰科的聚类结果一致,支持传统形态学分类结果。其中WCGS系统发育树自展支持率最高,更适合用于揭示物种的进化演变过程。WCGS构建的系统发育树,发现鹅掌楸属与木兰亚科植物亲缘关系更为紧密;濒危单种属物种华盖木与云南拟单性木兰亲缘关系紧密;而馨香木兰构成了远离木兰亚科其他物种的一个独立分支,显示与其他木兰亚科物种相对较远的亲缘关系。本研究明确了适用于科属间的系统发育分析方法,在叶绿体DNA分子层面构建了木兰科的系统发育关系,为该科物种的精准鉴定、种质资源开发、濒危物种引种保护等提供科学依据。
猜你喜欢
作物学报(2022年2期)2022-11-06
社会科学战线(2022年7期)2022-08-26
贵州林业科技(2022年2期)2022-06-22
武夷科学(2019年2期)2019-12-20
新青年(2019年8期)2019-08-22
天然产物研究与开发(2018年10期)2018-11-06
食品与健康(2018年8期)2018-08-23
中成药(2018年3期)2018-05-07
创作评谭(2014年3期)2014-05-12
中学生英语高中综合天地(2008年10期)2008-12-22
