
山药种质资源SSR分析及初级核心种质库的构建
2022-06-16 09:37张艳芳王锦华赵令敏张晓蒙霍秀文
北方农业学报 2022年2期
杨 帆,张艳芳,王锦华,赵 圆,赵令敏,张晓蒙,张 勇,霍秀文
(1.内蒙古农业大学园艺与植物保护学院,内蒙古呼和浩特 010011;2.乌兰察布市农畜产品质量安全中心,内蒙古集宁 012000;3.巴彦淖尔市农牧业科学研究院,内蒙古临河 015000)
山药(Dioscorea oppositaThunb.)是具有双子叶植物特征的单子叶植物,为垂直向地生长的长圆柱形块茎。山药是典型的药食同源性蔬菜,是食用蔬菜中的上品,也是最早列入《本草纲目》的中药材之一[1-2]。山药味甘性温,有健脾和胃、生津益肺、美容养颜、增强抵抗力等功效[3]。山药因其极高的食用和商业价值在国内外市场备受欢迎。我国是山药主要的起源中心之一,山药种质资源根据生态区域和生物学性状被分为南、北方山药群(以长江流域为界)[4-5]。近年来,山药实施“南薯北移”的种植策略,南方优质山药因气候不同、不易贮藏等问题在北方难以完全表现其特点,甚至和北方山药野生种形态相近。内蒙古西部地区为我国山药种植的新兴重要产区,近年来,种质资源广泛交换,导致品种名称混乱,给山药新品种的选育带来挑战[6]。
基于遗传标记对作物遗传多样性和核心种质的研究较多,但多数是研究基因的间接表达,如形态学、细胞学和生化标记,且易受环境等外在因素影响,难以精确分析[7]。分子标记是在DNA 水平上对于生物体遗传结构的直接反映[8],与前三类方法相比稳定性好且不易受环境限制,结果准确[9]。彭斌[10]利用SRAP 分子标记对薯蓣及其5 种近缘种研究发现,野生薯蓣属物种间存在有性繁殖的方式,并有非常丰富的基因流。郭文等[11]利用AFLP 标记技术构建了22 份薯蓣属材料的DNA 指纹图谱,为薯蓣属植物品种改良提供了理论依据。KEDRA 等[12]利用ISSR 标记对来自埃塞俄比亚的70 份山药的遗传多样性作出了评估,揭示了山药在属水平上的高度变异。随着种质资源的大量收集,资源规模的扩大使保存和分析工作成为难题。遗传多样性代表了植物对环境的适应力[13]。核心种质是以最少数量的遗传资源最大限度地代表整个群体的遗传多样性,通过对部分代表性种质的优先评价,充分利用其中的优良基因,可以大大提高种质资源的保存和利用效率[14]。SSR 标记是分子标记中一种简单快速、易于分析、相对稳定的方法。目前,国内外利用SSR分子标记技术展开了很多研究,崔洪宇等[15]、王郅琪等[16]、李琼等[17]、DUAN 等[18]、SIMONE 等[19]分别基于SSR分子标记技术分析了黄瓜、芝麻、大豆、马铃薯、番茄的遗传多样性,并将该技术用于品种鉴定并构建核心种质库。本试验利用SSR分子标记技术,分析来源于我国各地的55 份山药种质资源的遗传多样性,并基于聚类分析结果建立了山药初级核心种质库,旨在为山药种质资源保存和利用提供理论依据。
1 材料和方法
1.1 材料
供试55 份材料是近年来从我国山药主产区引进的种质资源,种植于内蒙古农业大学山药种质资源圃(表1)。在2021年8月取山药种质资源圃生长旺盛的山药细嫩叶片,立即将提前标记好的密封袋放入冰盒中,带回实验室储存到-80 ℃冰箱备用。

表1 供试山药材料编号及来源
1.2 DNA 提取及检测
根据试剂盒(天根生化有限公司)说明书中的步骤,提取55 份山药叶片基因组DNA。用1%琼脂糖凝胶电泳,紫外分光光度计检测DNA 纯度和完整性后,将OD260nm/OD280nm1.7~1.9 的DNA 稀释,并储存到-20 ℃备用。
1.3 SSR分析
引物来源:在内蒙古农业大学蔬菜育种课题组前期高通量测序获得的山药转录组分析结果中筛选40 组SSR 引物,由生工生物工程(上海)股份有限公司进行引物合成。经多次预试验,筛选出条带清晰、多态性高的12 组引物。
采用筛选出的12 对特异性好的SSR 引物,以55 份山药种质DNA 为模板进行PCR 扩增。PCR 反应体系:每20 μL 体系中含10 μL rTaq 酶、Forward primer 0.5 μL、Reverse primer 0.5 μL、DNA 模板0.4 μL、超纯水8.6 μL。反应程序:94 ℃预变性4 min;94 ℃变性30 s、54~60 ℃复性30 s(退火温度依据引物而定),72 ℃延伸1 min、循环40 次;72 ℃延伸10 min。产物于8%非变性聚丙烯酰胺凝胶中175 V电压下电泳2 h 左右,最后用硝酸银染色并拍照。
1.4 数据分析
根据试验所得电泳图,选择清晰、稳定的条带,在相同迁移位置上,统计扩增条带,有条带记为“1”,没有或者模糊条带记为“0”,得到“0/1”矩阵,使用POPGENE32 软件分析多态位点比率、平均等位基因数(Na)、平均有效等位基因数(Ne)、平均Nei′s 基因多样性指数(He)、平均Shannon-Wiener 多样性信息指数(I)。使用NTsys2.10e 软件中的UPGMA 方法进行聚类分析,用Tree plot 功能绘制聚类图,将数据用DCENTER 中心化后,采用EIGEN 系统进行主坐标分析[20]。
1.5 初级核心种质库的构建及评价
对种质资源进行聚类分析后,依据聚类图中的遗传相似系数采用最小距离逐步抽样法(LDSS)构建初级核心种质库,抽样比例为70%、50%、40%、30%、20%、10%,共抽样6 次[21]。具体方法为观察全部种质的UPGMA 聚类图,随机删除1 份遗传相似系数较大的种质组合,将剩余的种质再次聚类分析,以此类推,直到筛选出对于原始群体保留率较高的核心种质。每次抽样选出的种质,运用NTsys2.10e 软件对其进行聚类分析;下一次抽样在该聚类分析的基础上进行,以此类推。每次抽样后,均采用POPGENE32 软件分别对构建的初级核心种质进行遗传多样性分析,从而评价初级核心种质的代表性[22]。
2 结果与分析
2.1 山药种质的SSR 多态性分析
由图1 可知,采用筛选出的12 对多态性高、条带清晰、重复性好的SSR 引物,分别对55 份山药材料进行PCR 扩增,所扩增片段大多集中于250~750 bp。

图1 引物SSR-214 对55 份山药材料的PCR 扩增电泳图谱
由表2 可知,12 对引物扩增条带数为6~25,平均每对引物扩增出10.70 个条带,多态性条带数为6~24,平均10.50。12 对引物共扩增出128 个位点,多态性位点为126 个,多态性比率占98.59%,表明本试验检测的55 份山药资源遗传多样性十分丰富,种质有一定的代表性,可用于后续分析。

表2 供试SSR 引物及其扩增的多态性
2.2 SSR 标记山药种质资源遗传多样性分析
由表3 可知,经POPGENE32 软件分析得到55 份试验材料的遗传多样性指数,平均有效等位基因数(Na)为1.984 4,单个位点平均有效等位基因数(Ne)为1.420 4,有效等位基因比例为71.58%,平均Nei′s 基因多样性指数(He)为0.265 0,单个样品扩增条带平均Shannon-Wiener 多样性信息指数(I)为0.417 2。根据POPGENE32 软件分析结果可知,55 份山药材料的遗传相似系数在0.570~0.898,遗传距离在0.107 1~0.548 0。

表3 55 份试验材料的遗传多样性指数
2.3 山药SSR 标记聚类分析
采用NTsys2.10e 软件中的UPGMA 法,对SSR引物扩增出的128 条多态性条带进行聚类分析,得到聚类分析图。由图2 可知,种质间的遗传相似系数为0.570~0.898,河北保定山药(22 号)和山东嘉祥细毛长山药-1(26 号)遗传相似系数相对较大,为0.898,表明其亲缘关系相近;而河南铁棍山药-6(14 号)和贵州安顺山药(43 号)、山东嘉祥细毛山药-1(13 号)
和贵州山药(47 号)遗传相似系数相对较小,为0.570,表明这些材料的亲缘关系较远。
由图2 可知,在遗传相似系数为0.69 处可将55 份山药种质分为两类,第1 类包括日本山药-1(1 号)、河北双胞山药(8 号)等49 份种质,占供试材料的89.09%。进一步在遗传相似系数为0.74 处可将第1 类大部分种质分为5 个亚类,第1 亚类包括日本山药-1(1 号)等27 份材料;第2 亚类包括江西南城淮山药(34 号)等7 份材料;第3 亚类包括甘肃山药(28 号)、山东嘉祥细毛山药-3(40 号)和河南淮山药(41 号);第4 亚类包括广东淮山药(44 号)和云南播乐赖皮山药(51 号);第5 亚类包括河南山药(11 号)等8 份材料。海口山药(55 号)与江西泰和山药(33 号)在第1 类中被单独聚类,没有明显的聚类关系。第2 类包括西施山药(5 号)、河南温县铁棍山药(17 号)、陕西铁棍山药(31 号)、河南铁棍山药-5(20 号)、山东嘉祥细毛山药-1(13 号)和河南铁棍山药-6(14 号)共6 份材料。
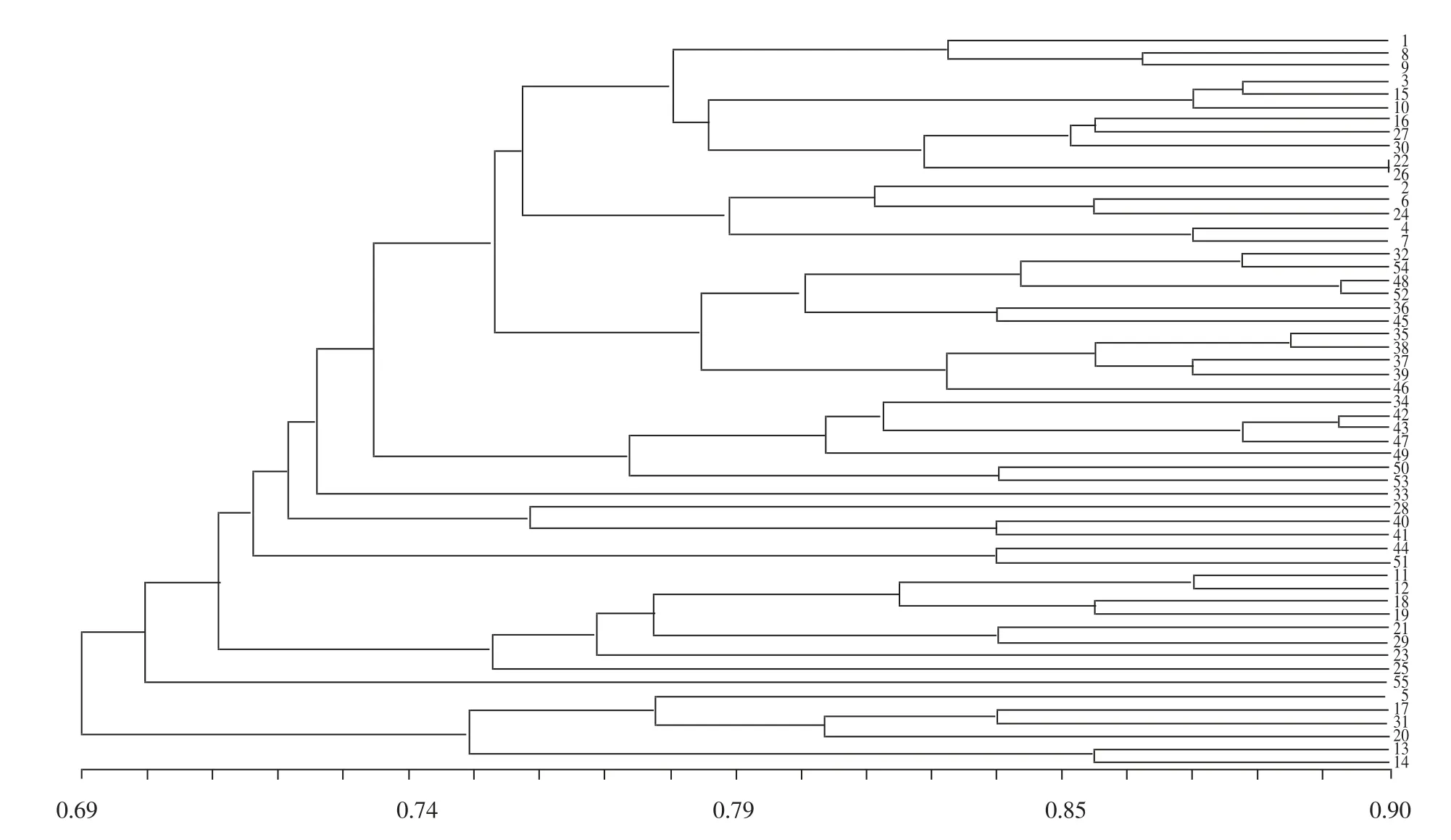
图2 55 份试验材料基于SSR 标记的聚类分析
2.4 山药初级核心种质库的构建
通过观察55 份山药材料的UPGMA 聚类图,用最小距离逐步抽样法(LDSS)构建核心种质库,抽样比例为70%、50%、40%、30%、20%、10%,共抽样6 次。每次抽样后构建的山药核心种质遗传多样性情况见表4。
由表4 可知,每次抽样后遗传多样性参数虽然总体变化不大,但各个指标均有变化。随着抽样比例的减少,等位基因数目(Na)不断减少,在第4 次(抽样比率为30%)抽样后明显减少。有效等位基因数目(Ne)在6 次抽样中不断缓慢增长。Nei′s 基因多样性指数(He)在前5 次抽样中均有少许增长,在第6 次抽样时明显减少。Shannon-Wiener 多样性信息指数(I)缓慢增长,在第4 次抽样后达到最高值,在第5 次抽样(抽样比例为20%)后出现下降的趋势,第6 次(抽样比例为10%)明显下降。多态位点和多态位点率从第2 次(抽样比例为50%)抽样后呈现下降趋势。经过以上分析,第4 次和第5 次抽样材料比其他抽样材料更有代表性,虽然在第5 次抽样时有效等位基因数目和Nei′s 基因多样性指数均大于第4 次抽样,但在第4 次抽样时等位基因数目、Shannon-Wiener 多样性信息指数、多态位点和多态位点率与第5 次相比更接近原始群体,且本试验所选山药材料不能涵盖全部,所以核心种质库数目太少将降低对原始群体的代表性。综上所述,确定抽样比例为30%时构建的种质库为SSR分子标记的初级核心种质库,图3 为本试验构建的基于SSR分子标记的山药初级核心种质聚类图。

表4 山药初级核心种质遗传多样性

图3 抽样4 次构建的山药初级核心种质SSR分子标记聚类图
2.5 主坐标分析
利用NTsys2.10e 软件对构建的山药初级核心种质库和留存的备用种质进行主坐标分析,验证其代表性。由图4 可知,55 份山药材料初级核心种质在原始群体中分布较分散,基本没有重叠现象,说明该初级核心种质有良好的代表性,可以代表原始群体的遗传多样性。

图4 山药初级核心种质与留存种质的主坐标分布图
3 讨论与结论
SSR分子标记因检出率高、操作简便、引物通用性强等优点已在植物遗传多样性等方面被广泛应用。高玉坤等[23]利用SSR分子标记和表型性状分析了65 个马铃薯品种遗传多样性,发现表型性状容易受环境因素和人工识别误差影响,与之相比SSR 标记的结果更为精确。而张燕梅等[24]用剑麻SSR 引物成功在中美麻和丝兰麻等近缘属中扩增出条带,证明SSR 引物具有较高通用性。SSR分子标记研究山药遗传多样性是可行的。本试验利用12 对SSR 引物对55 份山药种质DNA 扩增条带进行遗传多样性分析,共扩增出多态性条带126 条,多态位点率为98.43%,表明山药种质在分子水平上具有丰富的多态性。
本试验供试山药材料的128 条扩增条带平均有效等位基因数为1.420 4,有效等位基因比例为71.58%,Nei′s 基因多样性指数为0.265 0,Shannon-Wiener 多样性信息指数为0.417 2,遗传相似系数为0.570~0.898。上述指标表明了55 份山药材料丰富的遗传多样性。通过聚类分析,河北保定山药(22 号)和山东嘉祥细毛长山药-1(26 号)亲缘关系相对较近,遗传相似系数为0.898,从表型性状上看二者存在一定差别,但在一些优良特性上可能有相同的遗传特性而被种植户和学者广泛引种,这与陈占勇等[25]按山药花粉形态分类得到的结果一致。河南铁棍山药-6(14 号)和贵州安顺山药(43 号)、山东嘉祥细毛山药-1(13 号)和贵州山药(47 号)遗传相似度最小,亲缘关系相对较远。本试验在遗传相似系数为0.69 处可将55 份山药种质分为两大类群,第1 类包括日本山药-1(1 号)等49 份材料,第2 类包括西施山药(5 号)等6 份材料。在遗传相似系数为0.74 处可进一步将第1 类群大部分种质分为5 个亚类。海口山药(55 号)与江西泰和山药(33 号)在第1 类群中没有明显聚类关系,分别自成一类。依据田间观察的形态特征[26],本试验供试山药材料中52 份种质可归于普通山药,剩余第1 类群中的海口山药(55 号)和第4 亚类的广东淮山药(44 号)、云南播乐赖皮山药(51 号)应属于田薯,说明形态特征上相似的3 份田薯材料在其SSR分子水平上有一定的差异,而广东淮山药和云南播乐赖皮山药表现出较高的遗传相似度(0.840),但其在形态特征上存在差异,这与刘向宇等[21]利用ISSR分子标记方法对山药种质资源聚类分析结果相似。本试验所选的3 份山东嘉祥细毛山药和6 份河南铁棍山药虽然都各属于同一种,但在分子水平上显现出了一定差异,山东嘉祥细毛山药-1(13 号)与山东嘉祥细毛山药-3(40 号)亲缘关系相对较远,遗传相似系数为0.594,而与河南铁棍山药-6(14 号)聚为一类,相似程度较高;河南铁棍山药-6(14 号)与河南铁棍山药-3(18 号)亲缘关系相对较远,遗传相似系数同为0.625。从本试验构建的初级核心种质聚类结果(图5)看,日本山药-1(1 号)与河北麻山药-2(30 号)、内蒙古罗家营山药(29 号)与河北安平白山药(36 号)亲缘关系相近,但并未根据来源地聚为一类;西施山药(5 号)与海口山药(55 号)分别单独聚类,为较特殊种质。上述情况可能是3 种原因造成的:由于山药材料栽植环境不同,为适应环境其基因发生突变导致个体之间出现了差异;不同地区频繁品种交流导致品种间出现混杂现象;也可能是本试验采用的SSR分子标记较少,未能反映较完整的情况,具体原因有待研究。
构建核心种质库是目前研究种质资源的重点,李自超等[27]研究认为,核心种质应是动态变化的,取样策略很关键,应据具体物种而定。本试验采用最小距离逐步抽样法构建核心种质。观察聚类分析图多次抽样并分析其遗传多样性,发现除多态性比率较抽样前明显降低外,其余指标变化较小,Nei′s 基因多样性指数(He)和Shannon-Wiener 多样性信息指数(I)在第5 次和第4 次抽样后达到最高值。第4 次抽样的比率为30%,Nei′s 基因多样性指数、Shannon-Wiener 多样性信息指数分别为0.278 7 和0.433 1,高于Nybom 基于RAPD 和ISSR 等显性标记统计的多种植物种群水平的遗传多样性平均值(0.22)[28],且多态位点比率和等位基因数的保留率均达到抽样前的90%以上,符合核心种质资源代表性为70%~80%的要求[29],综合种质库数目分析后最终确定抽样4 次为初级核心种质,包含17 份山药材料,海口山药(55 号)、西施山药(5 号)可在今后的山药育种工作中作为重要种质加以利用。同时本试验构建的初级核心种质可为山药种质资源的管理、利用以及新品种的选育提供指导。
猜你喜欢
广西植物(2022年8期)2022-09-07
渔业科学进展(2022年4期)2022-09-05
农业工程学报(2022年8期)2022-08-08
中国农学通报(2022年12期)2022-06-01
自然灾害学报(2022年2期)2022-05-10
中国典型病例大全(2022年9期)2022-04-19
作物学报(2022年4期)2022-02-10
中国学校体育(2021年10期)2021-04-26
中学生物学(2019年7期)2019-10-17
中国当代医药(2009年22期)2009-05-14
