
黄土丘陵沟壑区不同草灌植被土壤分离速率及其主导因素
2022-06-15 14:33尚永泽李占斌马建业王晨光
水土保持通报 2022年2期
尚永泽, 马 波,2, 李占斌,2, 马建业, 王晨光
(1.西北农林科技大学 水土保持研究所 黄土高原土壤侵蚀与旱地农业国家 重点实验室, 陕西 杨凌 712100; 2.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100)
黄土丘陵沟壑区由于地理位置的过渡性、地形和地貌的复杂性、土壤的易蚀性及人类对土地的过度利用,多年来一直受到国内外学者的高度关注,同时也是国家退耕还林还草及生态建设的重点区域[1]。大规模生态建设使得该区土地利用结构发生了深刻变化,区域生态环境得到了明显改善,而土地利用方式的扭转进一步改变了土壤抵御水蚀的能力。现有研究表明,随着演替的进行,该区逐步由一年生杂草类群落发展为根茎禾草群落或多年生草本群落,并最终形成以小灌木+多年生草本群落为主的群落模式[2],曾光[3]通过对不同土地利用下的土壤进行冲刷试验发现,灌木林地和草地具有较强的抗冲能力,同时管世烽[4]通过对纯灌、纯草和草灌恢复模式下进行抗蚀抗冲试验,发现草灌混合模式下的土壤抗水蚀效果突出。草灌混生模式已经成为黄土高原坡面植被恢复的主要形式且对水土流失的治理有重要作用,因此,研究不同草灌搭配形式的生态抗蚀效以及对该区域生态建设效果的评估具有重要意义。土壤分离是指在降雨击溅或径流冲刷作用下,土壤颗粒脱离土体、发生位移的过程[5],是土壤侵蚀的起始阶段,探究土壤分离对于认识侵蚀过程与机理具有重要作用。已有研究表明,植被的地下根系是植物发挥抗水蚀作用最主要的部分[6-7],其主要通过物理固结和生物化学固结两方面的作用实现削减土壤分离的目的[8],且以物理固结为主导[9],李强[10]指出根系的物理固结作用约占根系抗蚀作用的77.7%~82.0%,是根系抗蚀的主要作用方式。根系构型也会影响根系固结作用的发挥,有学者指出,直根系和须根系植物在降低土壤分离能力的作用有明显差别[8,11],须根系植物的根表面积较大,能够与土壤颗粒充分接触,将更多的土壤颗粒固结在根系周围,其对土壤分离能力的削减作用较直根系高24.1%[12]。由于根系构型的不同,使得根系的减蚀效应随植物种的差异而显著不同。目前,国内外学者针对根系影响土壤抗侵蚀的研究多集中于单一特定植物种类,对于复合根系的土壤减蚀效果研究较少,虽然有部分学者开展了关于草灌复合根系的减蚀效应研究,但是由于野外自然环境不可控、采样取土困难等因素,野外原状土试验较少。为使研究更接近自然条件下根土侵蚀状况,本文选择黄土高原北部分布较为广泛的柠条锦鸡儿(Caraganakorshinskii)和冰草(Agropyroncristatum)、柠条锦鸡儿和铁杆蒿(Artemisiagmelinii)混生群落为研究对象,并以单一柠条锦鸡儿、冰草和铁杆蒿样地为对照,探究复合根系的减蚀效应,并分析影响其分离过程的主控因子,以期为黄土高原生态建设效果的评估提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于黄土丘陵沟壑区陕西省绥德县韭园沟中游左岸的王茂沟流域,面积5.97 km2,主沟长3.75 km,沟壑密度4.31 km/km2。该流域地表覆盖马兰黄土,厚20~150 m。受季风强烈影响,雨量少而分布不均,多年平均降水量 513 mm,多集中在汛期(6—9月),约占全年降水的73.12%,多以暴雨形式出现,一次暴雨产沙量往往为全年总产沙量的60%以上[13]。该区土质贫瘠,地块破碎,地貌复杂,在黄土丘陵沟壑区第一副区具有一定代表性。研究区灌木林主要以柠条锦鸡儿、沙棘(Hippophaerhamnoides)、达乌里胡枝子(Lespedezasdavurica)为主;撂荒草地主要植物群落有:冰草、铁杆蒿、白羊草(Bothriochloaischaemum)等。其中柠条锦鸡儿、冰草及铁杆蒿属优势群落,且在该区有广泛分布,具有一定代表性。
1.2 研究方法
1.2.1 样品采集 在2020年9月,选择以柠条锦鸡儿(下文简称为锦鸡儿)、冰草、铁杆蒿、柠条锦鸡儿+冰草和柠条锦鸡儿+铁杆蒿为代表的植物群落,且无明显人为扰动痕迹,植被长势均匀的5个样地作为研究对象(表1)。利用直径为10 cm,高5 cm的特制圆形取样器采集原状土样,每个样地均匀布设4个采样点。在冰草和铁杆蒿样地中,为防止丛生基部根系过多而影响取样器的打取而造成的土壤扰动,选择避开草本基部选择基部四周作为样点,由于样地具有较高的盖度,取样器内总是包含完整植株且距基部距离较近,具有一定代表性。用剪刀剪除地上部分,去除结皮及枯落物,最大程度剔除外源输入物质对分离能力的干扰。为尽量减少采样过程中扰动,须在采样点周围提前开挖深度15 cm左右的土壤剖面,将取样器垂直砸入地面,当环刀口与地面齐平时,对周围的土壤和根系进行挖掘和切割,以保证对样品的干扰最小,仔细取出样品,并对环刀下端的含根土体进行削剪。对于灌木样地,选择林下无草本生长,且生长状况相近的灌木植株作为研究对象,由于灌木根系在0—10 cm深度内的分布较为不均,因此在距离灌木基部50 cm处[14]挖掘20 cm深的剖面,简单确定灌木根系的走向,以保证取样器中含有灌木根系。去除土壤表面的结皮,将取样器垂直压入地面,后续采样方法如草本植物样地。对于草灌复合样地,选择灌木生长状况较为相近,林下草本植物较为单一,且与草本植物样地所选盖度相似的区域作为研究样点,依据灌木样地采样点确定方法,去除草本植物地上部分及结皮和枯落物后,采样方法与草本样地的采样方法一致。采集深度为0—10 cm,每个样点分0—5 cm和5—10 cm共采集4个土壤分离样品作为重复,每个样地共采集16个分离样品。本次试验共计采集80个分离样品。

表1 不同草灌植被下的样地概况
1.2.2 坡面放水冲刷试验及分离速率计算 本试验于2020年10月进行。冲刷槽长4 m,宽0.15 m,其中试验段为直径10 cm,深5 cm的圆柱体卡槽,位于冲刷槽中间且距冲刷段尾部30 cm。根据当地暴雨水平下平均次降雨量设计流量为36 L/min,坡度为15°。将清漆均匀的涂抹在土槽表面以固化床面,随后将直径小于5 mm的土壤颗粒均匀的洒在覆盖有清漆的土槽表面,再次喷洒清漆,使其包裹住表面的土壤颗粒。被固定的土壤颗粒可以在床面形成稳定的糙率,且在试验中水流无法改变坡面糙率;这样模拟天然土壤地表糙率,可以最大程度消除下垫面糙率变化对水流阻力等的影响[15]。样品采回后在水中浸泡12 h,使待测样品完全饱和,取出后在铁架上放置12 h,除去重力水,冲刷开始时用秒表计时,并在过程中利用测针观测土样的被冲刷深度,当冲刷最深处达2 cm时便停止冲刷,冲刷结束后将泥沙放入烘箱中烘干,记录干土重。
试验中径流水流分离能力为单位时间单位面积内径流分离土壤的质量即土壤分离。计算公式为:
(1)
式中:Dr为土壤分离速率〔kg/(m2·s)〕;W为试验过程中收集的泥沙样品干重(kg);t为水流冲刷时间(s);A为取样器投影的面积(m2)。
1.2.3 测定方法及数据处理 土壤理化性质及根系特征的测定于2020年11月进行。土壤黏结力(COH, kg/cm2)采用14-10型袖珍剪切仪进行现场测量[16],土壤容重(BD, g/cm3)采用环刀法测量[17],>0.25 mm水稳性团聚体含量(R>0.25, %)测定采用湿筛法[18],土壤黏粒含量(CLAY, %)采用马尔文激光粒度仪测定。
将冲刷后的取样器内的土体置于0.25 mm的土壤筛上反复冲洗得到根样品,而后采用扫描仪在400 dpi下进行彩色扫描,利用Win RHIZO 2009根系分析系统,对根长度,根表面积进行统计分析,扫描后的根系通过烘干法获得根干重。通过计算得到根重密度(RMD, mg/cm3),根表面积密度(RSD, 10-2m2/m3)和根长密度(RLD, cm/cm3),计算公式为:
(2)
(3)
(4)
式中:RM为根系总重(mg); RSA为根系表面积(m2); RL为根系长度(cm);V为取样器体积(cm3)。
将相关试验数据Microsoft Excel 2010中进行初步整理和表格的绘制,通过SPSS 26.0进行数据的差异性显著性检验(LSD法,p<0.05)和双变量相关分析(Pearson,p<0.05),通过Origin 2018进行相关性热图绘制。
2 结果与分析
2.1 不同草灌复合下的土壤分离速率
不同草灌复合类型的土壤分离速率的变化如图1所示。不同类型下土壤分离速率依次为:铁杆蒿〔0.200 kg/(m2·s)〕>锦鸡儿〔0.182 kg/(m2·s)〕>冰草〔0.063 kg/(m2·s)〕>锦鸡儿+铁杆蒿〔0.050 kg/(m2·s)〕>锦鸡儿+冰草〔0.028 kg/(m2·s)〕。相对于锦鸡儿、冰草和铁杆蒿等单一群落结构下的根系,草灌混生下的平均土壤分离速率降低了73.58%。其中,直根系的锦鸡儿和须根系冰草在复合后的分离能力最低,相较于锦鸡儿降低了84.65%,较冰草降低了55.37%,且较直根系组合锦鸡儿+铁杆蒿的分离速率低44.38%。在几种植被类型中,草灌复合条件下的土壤分离速率最低,其中以锦鸡儿+铁杆蒿为代表的直根系与直根系植物组合后的土壤分离速率减小的最为明显,而以锦鸡儿+冰草为代表的直根系与须根系组合的削减效果最佳。

注:JJ为柠条锦鸡儿,H为铁杆蒿,BC为冰草,JH为柠条锦鸡儿+铁杆蒿,JBC为柠条锦鸡儿+冰草; 不同小写字母表示不同复合类型间差异显著(p<0.05);下同
2.2 不同草灌复合下根系及土壤形状变化特征
表2为不同草灌复合下的根系特征和土壤性质。其中两种复合根系条件下的RLD,RSD和RMD分别较单一根系下的平均水平提高了22.58%,23.49%和31.05%。其中锦鸡儿+铁杆蒿的RSD较锦鸡儿显著增加了55.04%,锦鸡儿+冰草的RLD,RSD和RMD较锦鸡儿也有显著增加,分别增加了44.32%,62.79%和88.30%。另外锦鸡儿+冰草的RMD,RSD和RLD较锦鸡儿+铁杆蒿高19.59%,5.00%和10.49%。这说明草灌混生后根系密度明显增加,其中锦鸡儿+冰草组合下的根系密度最高。对于锦鸡儿+铁杆蒿和锦鸡儿+冰草两种复合根系而言,COH,R>0.25的平均水平分别较单一灌木及草本根系提高了35.15%和96.23%。其中锦鸡儿+铁杆蒿的COH分别较锦鸡儿和铁杆蒿分别显著提高了37.68%和58.33%,BD则与锦鸡儿处于同一水平,但较铁杆蒿显著增加,增幅达12.96%。R>0.25较两种单一植物均有增加,其中较铁杆蒿增加显著,达1.40倍,Clay较锦鸡儿数值较为接近,较铁杆蒿显著增加了30.26%。锦鸡儿+冰草的COH,BD和R>0.25较锦鸡儿和冰草均有所增加,其中COH分别增加了26.09%和19.18%,R>0.25分别提升了0.60倍和1.63倍。但是土壤黏粒含量(CLAY)较锦鸡儿增加了9.41%,较冰草减少了1.74%。这说明草灌混生后,COH,R>0.25增加明显。为探究不同植被模式下根系特征与土壤性质间的相互影响,对5种草灌复合条件下的根系和土壤因子进行相关性分析,结果详见表3。除锦鸡儿和铁杆蒿的根系指标分别与R>0.25和BD表现出较强的相关性,单一植物条件下的其他指标相关性并不显著,而复合条件下,锦鸡儿+铁杆蒿的RMD与COH,BD均显著相关,而锦鸡儿+冰草的RSD和RLD与BD呈极显著负相关,与CLAY呈显著正相关。说明根系构型的差别能有效影响土壤性质,这种表现在复合条件下表现的更为明显。
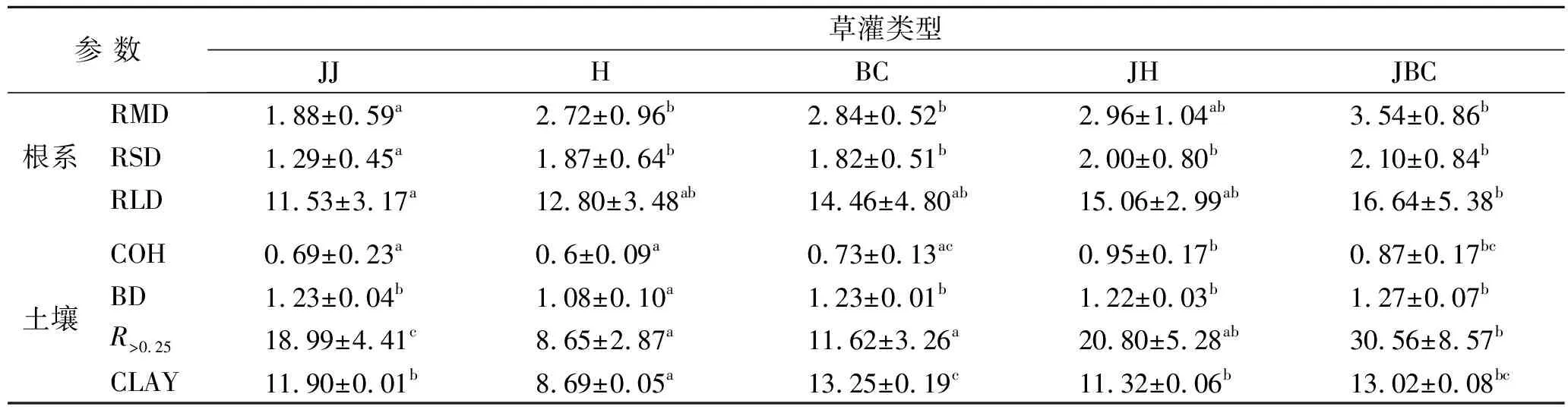
表2 不同草灌复合下的根系特征和土壤性状
2.3 土壤和根系指标对土壤分离速率的影响
由图2可知,根系指标中,与锦鸡儿和锦鸡儿+铁杆蒿的分离速率相关性最高的为RMD,与铁杆蒿和冰草相关性最高的为RLD,与锦鸡儿+冰草相关性最高的为RSD。土壤指标中,与冰草、铁杆蒿、锦鸡儿+铁杆蒿相关性最高的为R>0.25,与锦鸡儿+冰草相关性最高的为CLAY,其次为R>0.25,而与锦鸡儿相关性最高的为COH,其次为R>0.25,说明R>0.25是表征土壤分离速率的较优的土壤参数。此外,冰草和铁杆蒿的土壤分离速率与RSD,RLD等根系指标的相关性整体强于土壤性质的相关性。锦鸡儿、锦鸡儿+铁杆蒿和锦鸡儿+冰草的土壤分离率与根系的相关性接近于与土壤性质的相关性,说明在灌木和草灌混生条件下,土壤因子和根系因子共同作用影响了土壤分离速率。而在草地中,根系因子在提高土壤抵抗分离能力时发挥了更明显的作用。
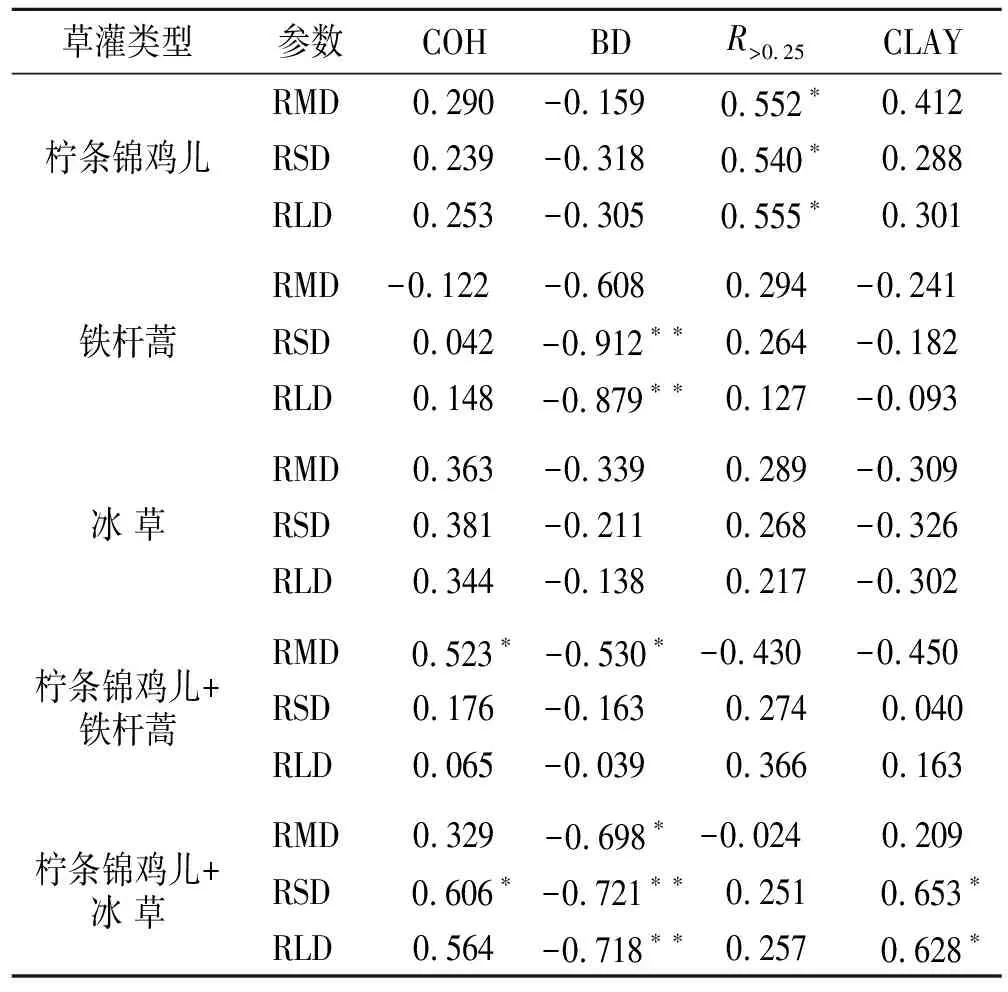
表3 不同草灌复合下根系因子和土壤因子的相关性
结合表3数据分析,两种复合模式下根系作用的发挥存在明显区别。锦鸡儿+冰草的RSD和RLD与土壤性质相关性更高且显著性更强,同时RSD与RLD同土壤分离速率相关程度也最高,说明RSD与RLD能够体现根系土壤性质的影响,相比于RMD在对土壤分离速率的解释上更具有优越性。而锦鸡儿+铁杆蒿的RMD不仅与土壤分离速率的相关程度最高,且与土壤指标的相关性更为显著,说明RMD是更重要的根系因子,而RSD与RLD则弱化为次要因子。
3 讨 论
本研究中,在两种组合根系条件下,土壤分离能力较单一植灌木和草本根系条件下的平均土壤分离率降低了73.58%,这与李强[8]在研究中柳枝稷和苜蓿的复合根系较单一根系显著降低的结论一致。相对于单一根系,草灌复合使得土壤中的根长密度和根重密度有效增加了18.42%和23.69%,而根系密度的增加使得土壤分离速率呈现明显的降低[19],因此复合根系下的土壤分离速率最小。根系密度的增加对土壤性质有明显的影响,一方面可通过增加分泌物的方式改善土壤团聚体、有机质和黏结力等特性[20,21],随着含根量的增加,土壤中的毛细根增多,根系在土体中交叉交织,固持土壤的作用随之增强,通过物理固结作用又可以增加土壤的黏结力[22]。更多的根系所分泌的各种类型的胶结物质固结了土壤[23],土壤性质的变化导致土壤可蚀性降低。同时,根系还可通过物理固结的方式,增加土壤团聚体含量,通过根的拌缠作用下截留黏粒使其胶结成大颗粒团聚体,由于大颗粒的水稳性团聚体遇水不易分散且具有较高的稳定性,其形成有利于增加土壤的抗侵蚀阻力,使土壤变得难以分离,进而削弱土壤分离率。另外,更致密的根系可以在土壤表面形成冲刷切口时,承担一部分的水流剪切力,以减小水流对土壤的冲击。然而,由于根系的穿插、死根的萎缩腐解使土壤孔隙度增加,进而降低了土壤容重,经过相关性分析可知,容重并非影响土壤分离速率的单一因素,还受诸多土壤和根系因素的影响,当其他因素对土壤分离能力的影响大于容重时,土壤分离速率并不会因为容重的减小而变化[24]。
研究结果表明,直根系和须根系复合下的锦鸡儿+冰草的分离能力较直根系与直根系复合下的锦鸡儿+铁杆蒿降低了79.81%,由于冰草为须根系草本,单位土体内的根系生物量较直根系的铁杆蒿有一定增加[25],其中冰草的根重密度和根长密度分别为铁杆蒿的1.13和1.04倍,更大的根系长度增加了根系与土壤的接触面积[26],使得其对土壤的固结能力更强。相关研究也表明,对于冰草和铁杆蒿而言,根系是决定其土壤分离速率最主要的因子[27]。但当其与锦鸡儿混生以后,不仅增加了单位土体的根系含量,改善了土壤性质,此外灌木较高的地表枯落物增加了土壤有机质,有机质的增加导致了团聚体含量的增加,有效削减了分离速率。这一现象在锦鸡儿、锦鸡儿+铁杆蒿和锦鸡儿+冰草中表现较为明显,锦鸡儿>0.25 mm水稳性团聚体的含量显著高于冰草和铁杆蒿。而锦鸡儿+铁杆蒿和锦鸡儿+冰草的>0.25 mm水稳性团聚体含量又相对于锦鸡儿有了提高。因此锦鸡儿+铁杆蒿和锦鸡儿+冰草的分离速率受土壤因子的影响显著增加。
由于两种复合类型的根系构型不同,根系因子的重要程度也有所区别。根表面积密度与根长密度是影响锦鸡儿+冰草土壤分离速率最主要的根系因子,根重密度则是影响锦鸡儿+铁杆蒿最重要的因子。以锦鸡儿+铁杆蒿为代表的直根系组合的根重密度与根长、根表面积密度相关性较低,说明根系生物量的增加并不能充分转化成与土壤界面的充分接触。而须根系植物在表层须根较多,根系在土壤中的穿插和缠绕能力较强,根—土接触面积更大且耦合也更为紧密,使得土壤因子与根系因子相互影响更为明显、剧烈,具体表现为对土壤容重、黏结力以及黏粒含量具有显著的的影响,使得其无论是在改善土壤性质和提高土壤抗蚀能力都更为突出,而直根系植物较须根系植物须根量大为减少,根土接触面积小,最终表现为根重密度的重要性提高,消减土壤分离能力的作用更弱。
本研究重点考虑两种不同根系构型条件下的草灌复合形式对土壤抗分离能力的影响,不同的根系构型使得根系形态特征具有明显差别,也对土壤产生了不同程度的影响,在根系和土壤的交互作用下进一步提高了根土复合体的抗冲性。探究不同的根系径级的分布和分泌物的生成对交互作用的体现具有一定价值,还需后续进一步探究。
4 结 论
(1) 在生态建设草灌逐渐趋于复合的过程中,土壤分离能力呈逐渐减小的趋势,表明草灌混生对提高土壤抗水蚀能力有积极作用,尤其以锦鸡儿和冰草为代表的直根系灌木和须根系草本的复合模式效果最佳,可以作为黄土高原地区一种有效的生态恢复模式。
(2) 复合条件下,根系特征与土壤性状间表现出了较强的相关性,根系密度与土壤黏结力、黏粒含量呈显著正相关,与土壤容重则呈显著负相关。影响土壤分离速率的主要根系因子是根重密度和根长密度,影响最大的土壤因子为>0.25 mm团聚体含量,草灌混生可以有效增加单位土体内的根系密度,增加土壤黏结力、>0.25 mm团聚体含量,其中以锦鸡儿+冰草为代表的直根系灌木与须根系草本的复合形式表现的最为明显。
(3) 草灌混生之后,由于根系因子对土壤因子的显著影响及团聚体含量的增加,使土壤因子对土壤分离速率具有更高的相关性。另外,两种混生模式的不同使根系发挥作用的因子有所不同,其中灌木与须根系草本的主导根系因子是根表面积密度与根长密度,而灌木与直根系草本主导根系因子是根重密度。
猜你喜欢
儿童时代·快乐苗苗(2022年7期)2022-10-18
贵州畜牧兽医(2022年3期)2022-06-28
童话王国·文学大师班(2022年2期)2022-02-05
内蒙古林业(2021年7期)2021-12-31
现代农村科技(2021年5期)2021-12-07
现代园艺(2021年23期)2021-12-01
乡村科技(2021年16期)2021-10-21
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
当代陕西(2019年7期)2019-04-25
