
低温对市政污水生物处理的影响及对策
2022-06-15 02:22:10张涛,余冉
净水技术 2022年6期
张 涛,余 冉
(东南大学能源与环境学院,江苏南京 210096)
活性污泥中的微生物种类/丰度和活性受季节变化影响大,以活性污泥为核心的污水生物处理工艺处理效率也随季节的变化而产生波动。尤其是在温度较低的冬季,活性污泥中的功能微生物会受到低温的生长抑制影响[1-2],出现微生物活性低、污泥膨胀、出水水质氮磷含量不达标等现象。研究表明,我国北方寒冷地区的污水生物处理工艺,在污水温度低于13 ℃时,整个污水处理系统处理效率会下降50%左右;而在水温低于4 ℃时,工艺的生物处理效果可忽略不计[3]。表1列举了我国北方城市污水处理厂不同污水处理工艺冬春两季的水温情况,冬春季的平均水温都低于15 ℃,最低水温甚至达到了4.7 ℃。因此,保障冬季低温市政污水处理系统稳定运行极为重要。
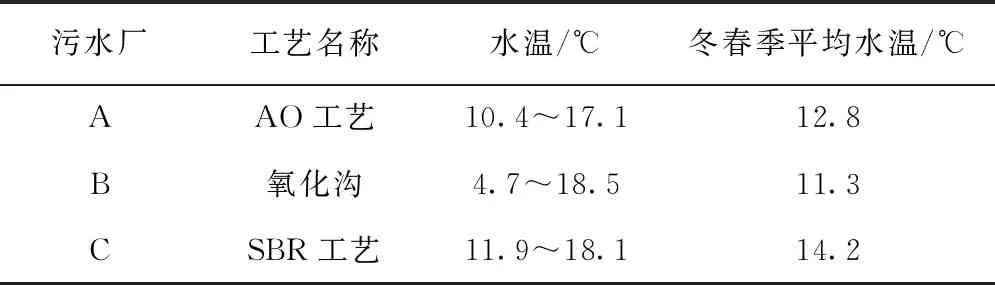
表1 我国北方地区不同污水处理工艺春冬两季水温[4]
1 低温对活性污泥的影响
1.1 活性污泥微生物
1.1.1 低温对污泥微生物群落结构的影响
活性污泥主要由微生物细胞组成,同时吸附了污水中的各种有机物和无机颗粒。活性污泥中的微生物种类丰富,一般以细菌为主,还含有古细菌、真菌、原生动物和后生动物等。它们能够形成一个完整的食物链,利用废水中的营养物质来维持群落结构的稳定。污泥处理工艺中,常见的优势菌门有变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、绿弯菌门(Chloroflexi)和放线菌门(Actinobacteria),以及许多未知菌种[5]。这些菌种在污泥中担任的角色各不相同,如硝化螺旋菌门(Nitrospirae)与活性污泥法的硝化作用有紧密关系[6],变形菌门和厚壁菌门(Firmicutes)与反硝化作用有关[7],以及与污泥膨胀相关的腐螺旋菌科(Saprospiraceae)、黄杆菌科(Flavobacteriaceae)、四球虫属(Tetrasphaera)[8-9]等。
温度是影响污泥微生物群落结构的重要因素。Ju等[1]在长达4年研究中,发现许多功能菌属丰度遵循季节变化规律,夏季硝化细菌——硝化螺旋菌属(Nitrospira)和亚硝化单胞菌属(Nitrosomonas)表现出更高的丰度,而冬季除磷菌——四球虫菌属(Tetrasphaera)、具有反硝化作用的副球菌属(Paracoccus)、潜在的人类粪便细菌双歧杆菌属(Bifidobacterium)、多利亚菌属(Dorea)和瘤胃球菌属(Ruminococcus)等的丰度表现得更高。季节变化在改变污泥微生物的种群丰度的同时可能会影响其功能基因的表达。Liu等[10]的研究发现,硝化单胞菌和硝化螺菌分别是代表性的氨氧化细菌(AOB)和亚硝酸盐氧化细菌(NOB),而这2种细菌的丰度在冬季下降并导致了氨氮去除率降低。同样,樊柳[11]对武汉市多个污水处理厂取样研究发现,同一水厂的暖季样品的Chao和Ace指数(Chao和Ace指数反映菌群丰度,数值越大,物种总数越大)高于冷季,而Simpson指数(Simpson指数反映种群多样性,Simpson指数值越小,表明物种越丰富)却具有一致性,这表明低温只会使得污泥微生物菌群丰度减小,而对污泥微生物种群多样性几乎没有影响。
污水处理工艺在冬季低温环境下经常发生污泥膨胀[12-13],污泥群落结构稳定性和污染物的去除效率都会受到负面影响。污泥膨胀一般分为污泥丝状菌膨胀和非丝状菌膨胀,其中非丝状菌膨胀又分为菌胶团污泥膨胀和菌胶团解体污泥膨胀。低温下的非丝状菌膨胀一般是菌胶团污泥膨胀,这种污泥膨胀可以通过改善活性污泥的组成,或调整适当回流比、溶解氧、混合液悬浮固体浓度(MLSS)等参数进行控制[14]。端正花等[9]对冬季郑州某污水处理厂发生的污泥膨胀现象进行了机理研究,利用高通量测序技术对1月—4月及12月发生污泥膨胀的污泥样进行分析,发现腐螺旋菌科和黄杆菌科的丝状菌在低温下的过度增殖是污泥膨胀的主要成因。高春娣等[15]发现在14 ℃的低温下SBR反应器的污泥丝状菌膨胀,污泥沉降性能明显恶化,污泥中特定菌群丝状菌群的丰度由0.49%变化至26.04%,而脱氮菌群和除磷菌群的丰度则相对减少。
1.1.2 低温对污泥微生物活性的影响
微生物的活性一般指微生物的新陈代谢的速率,即污泥微生物细胞与污水之间的能量和物质交换,以及污泥微生物细胞内物质和能量的转变。当环境温度降低时,微生物的新陈代谢速率会下降从而导致其活性降低。已有研究显示,低温对微生物活性的影响因素包括以下几点[16-22]。(1)膜的流动性。微生物细胞膜的基本结构是磷脂双分子层,这种结构需要保持适当的流动性,以保证基本膜蛋白的渗透性和活动。常温下的磷脂双分子层是液晶态的结构,而温度降低会使其向凝胶态转变,细胞膜因不能保持流动性而丧失物质交换功能。(2)维持低温下蛋白质的合成和产生以及维持酶催化反应的速率。低温条件下,分子内氢键增加导致酶的折叠增加,且单个酶分子合成减少,降低了酶催化反应速率,最终引起蛋白质合成速率的下降。(3)冷适应蛋白及其作用机制。低温微生物为了应对温度的突然降低,会在体内合成冷适应蛋白来适应温度的变化。(4)防冻机制。适应低温的微生物能够产生相容性溶质(甘氨酸、甜菜碱、甘油、海藻糖等),这些溶质能够在细胞质内积累,在不影响细胞活动的同时,使细胞免于低温伤害。
1.2 活性污泥理化性质
1.2.1 低温对活性污泥絮体结构的影响
活性污泥絮体的粒径分布、结构和密度是控制活性污泥固液分离性能的最重要物理参数。污泥絮体大、密度高将有助于污泥快速沉降,并且使泥水分离后上清液浑浊度低。污泥絮体结构受多种因素影响,如pH、Zeta电位、胞外聚合物(EPS)含量、丝状菌数量等。弱稳定的絮体结构遇到剪切力作用时,容易在活性污泥水处理过程中分解,导致污水中的细小颗粒和游离态细菌数量增加。进一步过程中,活性污泥不可避免地在水处理系统中各工段中会遇到因曝气、泵输送、脱水等产生的剪切力作用,污泥絮体往往会被分解成更小的颗粒,从而不利于泥水分离。而出水中30%的化学需氧量(COD)和50%~80%的磷都与粒径大于0.1 μm的颗粒相结合[23],低温对絮体的影响间接导致了出水水质下降。
冬季水温较低时,活性污泥絮体的絮体结构一般较弱,这使得污水中的悬浮固体浓度较高[24]。Koivuranta等[25]发现絮体的形成与进水水质没有明显关联,而受水温影响极大,具有明显的季节性特征。夏季的污泥絮体通常个体大,形状较圆,丝状菌数量少,且小颗粒数量较少,而冬季污泥絮体的以上相关参数与夏季相反。造成这一差异的原因可能是低温促进了丝状菌生长,致使污泥絮体中的丝状菌数量增加。包含大量丝状菌的絮体对剪切作用的敏感性更高,絮凝强度也降低,极易发生大规模破碎[26-27]。破碎后的絮体尺寸变小,同时小颗粒数目增加,导致出水水质变差。
1.2.2 低温对污泥絮体沉降性能的影响
污泥的沉降性能和松散程度一般用污泥容积指数(SVI)来衡量,过高的SVI会降低污泥沉降性能和脱水效果。污泥沉降性能的季节变化特性在污水处理厂中较为常见,SVI通常在冬季会远高于夏季,并且这一现象有时会伴随着污泥中丝状菌丰度的增加[28]。此外,冬季污水处理厂为弥补污水处理系统低温除磷的不足,经常会利用铁盐进行化学强化除磷。相关研究发现这一举措提高了污泥中的铁浓度,会对污泥沉降性能和压实性能产生负面影响[24]。
污泥的表面性质(疏水性、表面电荷)和EPS的组成也会影响污泥的生物絮凝性能,EPS的含量与污泥的沉降性能SVI呈正相关性[29-30]。EPS是细胞的代谢产物,主要由多糖、蛋白质、核酸等高分子物质构成,是污泥絮体中的第三大成分,仅次于细胞和水[31]。活性污泥组成会随着季节变化而变化,在冬季时受低温影响,污泥中的EPS含量达到全年最高[32-33]。崔迪[4]在对寒区污水处理系统的研究中发现,限制微生物合成多糖的限制酶EC 1.1.1.22(UDP-glucose 6-dehydrogenase,UGD)在冬季丰度达到最高,细菌利用多糖可以合成更多的EPS,这一定程度上解释了冬季污泥中EPS含量偏高的现象。
2 低温市政污水生物处理强化技术及运行调控策略
对于绝大部分市政污水生物处理工艺来说,低温会影响到其脱氮除磷效率但对COD的去除效率影响不大。故在《城镇污水处理厂污染物排放标准》(GB 18918—2002)中,当污水温度低于12 ℃时,对总氮和氨氮排放标准进行了适当放宽。
2.1 低温脱氮
传统的生物脱氮途径一般包括硝化和反硝化两个阶段,分别由硝化菌和反硝化菌主导完成。低温对脱氮的影响主要体现于对硝化和反硝化作用的抑制。常温硝化菌群的最适温度为30 ℃,世代时间约为15 h,而在低温5 ℃时则延长为200 h[34],故其硝化作用基本停止,而当温度低于4 ℃时,悬浮生长的污泥中可能不存在硝化菌[35-36]。同样,低温下的反硝化细菌,生长代谢速率降低,而且由于低温会使水中溶解氧(DO)增加,分子态的氧与硝酸盐竞争优先充当反硝化细菌的电子受体,抑制硝酸盐的还原,降低反硝化速率。
许多学者从环境中驯化筛选嗜冷菌,并验证它们在低温下处理污水的能力。盛晓琳等[37]在低温(10 ℃)下驯化低温硝化污泥,经过菌种富集之后,污泥中的硝化杆菌属和亚硝化单胞菌属的相对丰度提高了43倍和42倍,在面对低温影响时,该硝化污泥能够在5 d内,将氨氮去除率升至99%。孟盈盈[38]从北极海洋沉淀物中筛选出多株反硝化细菌构建混合菌群,发现其在10 ℃下,48 h内可将初始质量浓度为60 mg/L的氨氮完全去除。伍海全等[39]筛选驯化了嗜冷性微生物菌群并形成活性污泥处理水温为5、10、15 ℃的人工污水,结果表明COD去除率能够达到95%以上,氨氮去除率能够达到85%以上,且出水能够达到城镇污水处理厂污染物排放标准一级A标准。唐美珍等[40]从人工湿地的底泥中分离出黄假单胞菌,其能够在16 ℃下,对模拟生活废水中的COD、氮、磷达到较高的脱除效果。李晓亮[41]从北极海洋沉积物中分离出多株低温硝化细菌,利用其构建混合菌群在15 ℃下处理废水48 h能够获得91.7%的氨氮去除率。
研究表明,嗜冷氨氧化细菌在0~5 ℃时,氨氮的去除作用并不明显;而在8~10 ℃时,能够保持较高的硝化速率[42]。王思萌等[43]在以短程硝化反硝化-厌氧氨氧化一体化工艺(SPN/A)为主体的序批式活性污泥(SBR)反应器中,15 ℃的低温反应条件下定期投加短程硝化污泥,发现该方法能够增强AOB和厌氧氨氧化菌的丰度及活性,抑制NOB活性,从而增强系统的低温氨氮处理能力。
好氧颗粒污泥(AGS)技术具有沉降性能好、生物活性高、耐冲击能力强的特点。Jiang等[44]利用序批式气提反应器(SBAR)在低温(8 ℃)下快速造粒,系统运行10 d能够观察到紧致污泥颗粒,25 d能够稳定运行,该系统稳定后的氨氮去除率能够达到99%以上。
通过载体的利用可以固定富集嗜冷菌,从而保证污水生物处理系统中的低温菌生物量,避免因直接投加造成的菌株流失和菌株适应等问题,以提高污水在低温条件下的处理效能。郑志佳等[45]为解决移动床生物膜反应器工艺在冬季出水氨氮不达标问题,向生化池好氧区投加悬浮载体,低温条件下仍然能够保持硝化菌群数量,达到延长泥龄的目的,使得出水氨氮达标。
2.2 低温除磷
生物除磷需要保证充分的有机碳源。虽然低温对聚磷菌的生长速率影响不大,但是会降低大分子有机物的水解速率,导致易受聚磷菌降解的挥发性脂肪酸(VFAs)等小分子产物含量降低,从而间接减缓微生物好氧释磷作用。
在传统活性污泥法AAO工艺中,除磷的唯一方法是从二沉池排放剩余污泥,遇到温度降低时,工艺的除磷能力明显不足。为了增强传统AAO工艺的低温除磷效果,Li等[46]对AAO工艺进行了改进。他们在缺氧池和好氧池之间增加一个中沉池,专门进行硝化和释磷。并在二沉池后端连接一个厌氧释磷池,污泥释放磷后活性会表现得更高,将其回流至前端好氧池,能够更好地去除污染物,尤其是磷。与改进前相比,改进后的AAO工艺对磷的去除效率从56.0%增至93.3%。
虽然低温一般会导致磷去除率下降,但也有研究人员发现低温会促进磷的去除。Erdal等[47]在研究生物强化除磷系统(EBPR)时发现,水温由20 ℃降至10 ℃,生物除磷能力会下降,而降至5 ℃时,生物除磷能力反而会有所提高。Whang等[48]和Panswad等[49]也发现了类似的结论,低温会对除磷有一定的促进效果。他们认为这是由聚磷酸盐微生物(PAOs)和糖原累积微生物(GAOs)之间的相互竞争引起的,PAOs能够在低温下优先利用VFAs等有限碳源进行除磷,而当温度升高,GAOs会抢夺碳源抑制PAOs的生物除磷反应。
2.3 低温污水处理工艺的改造与运行调控
表2列举了多种污水处理工艺的低温改造和运行调控措施,工艺的提标改造措施除了优化原有工艺运行模式,多数都在原工艺的生物处理部分添加了填料,利用填料形成生物膜保留生物量的特性,增强低温环境下对氮、磷的去除能力。通过向生物反应池内投加载体实现污水降解效果提高的工艺有很多,代表性工艺有挪威的KMT工艺(将圆柱状KMT聚乙烯塑料载体与生物流化床相结合的污水处理技术)、美国的Captor工艺(利用聚氨酯泡沫为生物膜载体的污水处理技术)、Biofor工艺(利用曝气生物滤池内的填料附着生物膜的污水处理技术)、德国的Linpor工艺(利用聚氨酯泡沫块作为填料的生物流化床工艺,载体尺寸比Captor工艺的小)和IFAS工艺(活性污泥法与生物膜技术相结合和污水处理技术)等。其中,IFAS工艺在国内研究较为成熟,在北方污水处理中也发挥了显著效果[50-51]。

表2 低温污水处理工艺的改造与运行调控
3 总结
低温条件会导致市政污水生物处理系统氮、磷去除率下降,为克服现有问题,研究者们提出驯化筛选低温菌、工艺改良等技术研究,但目前仍然存在不足:(1)对嗜冷菌的研究多停留在小试试验基础,缺少中试及实际工程的应用研究;(2)对单一嗜冷菌的处理效果研究较多,而对复合菌种协同处理效果和机理的研究较少;(3)多数研究角度都是从污泥的物理性质和污泥组分的变化出发,而更深入的角度,如微生物酶、功能基因等研究较少。
与嗜冷菌的开发相比,对低温污水生物处理工艺的改进与调控策略的研究与工程应用成效更为显著。其中,基于生物膜技术的组合工艺因其能够在低温下保持较高的生物量和较好的处理效果,在未来的低温污水处理研究中可能更具有发展前景。
猜你喜欢
中央民族大学学报(自然科学版)(2018年3期)2018-11-09 01:16:42
环境保护与循环经济(2017年7期)2018-01-22 02:58:12
中央民族大学学报(自然科学版)(2018年3期)2018-01-12 06:03:03
计算机测量与控制(2017年6期)2017-07-01 16:24:28
中国环境科学(2016年3期)2016-02-08 15:07:14
哈尔滨工业大学学报(2015年2期)2015-09-21 01:41:04
化工管理(2014年9期)2014-12-11 05:45:58
应用海洋学学报(2014年2期)2014-11-26 01:20:46
应用化工(2014年1期)2014-08-16 13:34:08
深圳信息职业技术学院学报(2014年3期)2014-04-21 09:05:38
