
海洋天然产物壳寡糖抑制细菌生长和黏附的作用评价
2022-06-10 02:31温佳颖牛雪婷陈圣威陈宗周王新晨吴淑婷巨向红刘晓曦雍艳红广东海洋大学滨海农业学院动物医学系湛江524000广东海洋大学深圳研究院深圳518108
动物营养学报 2022年5期
温佳颖 牛雪婷 陈圣威 陈宗周 王新晨 吴淑婷 巨向红 刘晓曦 雍艳红*(1.广东海洋大学滨海农业学院动物医学系,湛江524000;2.广东海洋大学深圳研究院,深圳518108)
在水产品虾、蟹加工中,产生大量的下脚料虾、蟹壳,可提取甲壳素。甲壳素部分或全部脱乙酰基的产物为壳聚糖。壳聚糖可经化学降解或微波降解而形成壳寡糖(chitosan oligosaccharide, COS),它能生物降解,可溶性好,具有明显的抗菌[1]、收敛[2]、黏膜修复[3]、自由基清除和免疫增强活性。COS的毒性很低,只有高浓度的COS才会导致死亡,在小鼠口服急性毒性试验中最大耐受剂量超过10 g/kg BW,连续30 d用含3.0% COS的饲粮喂养大鼠也没有发现其异常症状和临床症状或死亡[4]。COS易于被肠道吸收,能有效缓解肠道炎症,并增强动物的免疫应答[5]。据报道,饲粮中添加COS可显著提高仔猪小肠绒毛长度,增加小肠绒毛高度与隐窝深度的比值[6],降低盲肠中大肠杆菌的含量和抑制金黄色葡萄球菌等细菌的生长[7]。由此可见,COS具有开发成“替抗”天然活性药物的潜力,用于解决动物养殖过程中日益严重的细菌耐药性和抗生素过量使用等问题。但COS的抑菌效果会受分子质量大小和添加剂量的影响[8],筛选并优化出特定分子质量的COS和适宜添加量具有重要意义。
黏附和定植是细菌入侵动物肠道的首要过程,而肠上皮的黏液层是阻止细菌入侵的重要保护屏障,由黏蛋白(MUC)2和MUC4等分子组成,肠道致病菌可通过不同机制降解MUC[9]。另外,大肠杆菌、沙门氏菌等致病菌可以通过调节细胞骨架蛋白、紧密连接蛋白和黏附分子,如闭锁蛋白(Occludin)、封闭蛋白-1(Claudin-1)及细胞间黏附分子-1(ICAM-1)而破坏黏膜屏障[10-11]。研究发现,COS有一定的抗黏附性,改性COS对皮肤革兰氏阳性菌和革兰氏阴性菌均表现出良好的抗菌黏附性能[12]。但有关COS对肠道致病菌的抗黏附作用及机制的研究不多。因此,本研究检测了COS对致病性大肠杆菌、肠炎沙门菌、金黄色葡萄球菌和福氏志贺菌的最低抑菌浓度(MIC)和最低杀菌浓度(MBC),筛选了COS对4种常见致病菌的最佳添加量,并初步探究了COS抑制致病菌黏附的分子机制,这为COS作为替抗产品的进一步开发和安全使用提供了试验及理论依据。
1 材料与方法
1.1 试验样品及来源
COS(分子质量<3 000 u,脱乙酰度>90%)购自上海某生物有限公司;BNCC337272致病性大肠杆菌购自北纳生物有限公司;CMCC51572福氏志贺菌、ATCC14028肠炎沙门氏菌和ATCC6538金黄色葡萄球菌购自上海保藏生物技术中心。猪肠上皮细胞IPEC-J2细胞购自北京北纳生物科技有限公司。
1.2 主要材料与试剂
试验主要试剂见1。
表1 主要试剂
1.3 试验方法
1.3.1 细胞培养
IPEC-J2细胞生长于含1%双抗、10%胎牛血清的DMEM/F12完全培养液中,在37 ℃、5% CO2的培养箱中培养。1~2 d更换细胞培养液,待细胞融合80%时使用0.25%含乙二胺四乙酸(EDTA)的胰酶消化进行传代。
1.3.2 细菌培养
储存菌液浓度约为1×108CFU/mL,用含有50%甘油的肉汤培养基储存细菌,并分装于冻存管置于-80 ℃超低温冰箱中保存,待用时取出冻存管,用接种环挑取少量菌液接种于高压灭菌的肉汤培养基中,培养18~24 h,观察细菌的生长状态,然后用接种环挑取菌液再接种于肉汤培养基中培养待用。
1.3.3 MIC和MBC的测定
采用试管二倍稀释法进行COS的MIC的测定,无菌操作条件下,取7支10 mL无菌试管,排序1~7号,将1~5号试管的COS浓度依次稀释为51.2、25.6、12.8、6.4、3.2、1.6 mg/mL,6号试管不加COS作为阳性对照,7号试管仅加入4 mL肉汤培养基作为阴性对照,接种环挑取菌液接种1~6号试管。重复3次。将各试管置于37 ℃培养箱中培养24 h,观察结果。培养基呈清澈,能够抑制80%细菌生长的最低药液浓度即为受试菌MIC。将培养后的含菌培养液混匀,取10 μL采用连续划线方法接种于营养琼脂平板。将培养皿置于37 ℃培养箱中培养24 h,观察细菌生长情况并记录菌落数。菌落数少于5个或降低3个数量级活菌的最低药液浓度即为受试菌MBC。
1.3.4 细菌黏附率的测定
采用一种基于羟基荧光素二醋酸盐琥珀酰亚胺脂(CFSE)荧光的方法检测细菌黏附性[13]。细菌中加入50 μmol/L CFSE,37 ℃孵育30 min,重悬于无菌磷酸盐缓冲液(PBS)中,调整细菌菌落数为1×108CFU备用。将IPEC-J2细胞接种于全黑96孔板(密度1×103个/孔)中,每组设6个重复孔。细胞完全贴壁后,设置空白组(CON组,无COS及细菌)、阳性对照组(无COS,加入细菌)及COS组(200、400、600和800 μg/mL COS),培养24 h。弃细胞上清液,每孔加入200 μL CFSE标记的受试菌菌液,培养1 h,PBS洗除未黏附的细菌。每孔加入200 μL含1%(w/v)十二烷基硫酸钠的0.1 mol/L NaOH溶液,在37 ℃孵育1 h,之后用移液枪吹打混匀。最后使用全波长荧光酶标仪在λex 485和538 nm条件下检测每孔细胞的荧光值。用空白组的荧光值校正阳性对照组和试验组的荧光值,黏附性表示为校正后的试验组荧光值与阳性对照组荧光值的百分比。
1.3.5 实时荧光定量PCR(RT-qPCR)
消化受试细胞,TransZol法提取细胞总RNA,gDNA Removal and cDNA Synthesis SuperMix试剂盒反转cDNA,RT-qPCR检测细胞ICAM-1、MUC2、MUC4、Occludin、Claudin-1和c-Jun氨基末端激酶(c-Jun-terminal kinase,JNK)的表达水平。RT-qPCR根据试剂盒说明书进行操作。表2列出了本研究中使用的引物序列,引物由生工生物工程(上海)股份有限公司合成。试验结果通过2-ΔΔCt方法计算基因相对表达量。

表2 实时荧光定量PCR引物
1.4 数据统计分析
试验数据用SPSS 26.0软件进行单因素方差分析,Tukey’s进行多重比较,使用Graphpad prism 8.0软件绘制图表。P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 COS对肠道致病菌的MIC和MBC
由表3可知,与对照组相比较,COS对致病性大肠杆菌的抑菌和杀菌效果较明显,MIC仅为1.6 mg/mL,MBC为12.8 mg/mL。COS对金黄色葡萄球菌、肠炎沙门菌、福氏志贺菌均具有抑菌作用,MIC为6.4 mg/mL,MBC大于25.6 mg/mL。
2.2 COS对肠道致病菌黏附IPEC-J2细胞的影响
IPEC-J2细胞加入不同浓度的COS后预孵育24 h,再与CFSE标记的细菌共培养1 h。结果表明,COS浓度与金黄色葡萄球菌、致病性大肠杆菌、肠炎沙门菌和福氏志贺菌的黏附率均存在显著非线性相关(图1-A~图1-D;R2>0.850 0),不同浓度COS均可极显著抑制该4种细菌对IPEC-J2细胞的黏附(图1-A~图1-D;P<0.01),其中600 μg/mL的COS对金黄色葡萄球菌黏附率的抑制作用较明显(图1-A),而800 μg/mL的COS对致病性大肠杆菌、肠炎沙门菌和福氏志贺菌黏附的抑制作用较明显(图1-B~图1-D)。
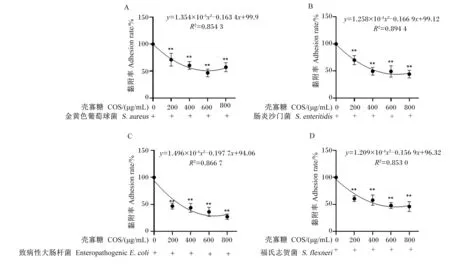
A~D:不同浓度COS预处理下金黄色葡萄球菌、肠炎沙门菌、致病性大肠杆菌和福氏志贺菌黏附IPEC-J2细胞的黏附率。** P<0.01; * P<0.05。
2.3 COS对肠道致病菌黏附IPEC-J2细胞后MUC基因表达的影响
致病性大肠杆菌、肠炎沙门菌和福氏志贺菌与IPEC-J2细胞共培养后,细胞中MUC2和MUC4基因相对表达量极显著降低(图2-A和图2-B;P<0.01),而COS预处理能极显著抑制肠炎沙门菌和致病性大肠杆菌侵染所致的MUC2和MUC4基因相对表达量的下调(图2-A和图2-B;P<0.01)。金黄色葡萄球菌侵染IPEC-J2细胞后,细胞中MUC4基因相对表达量极显著下降(图2-B;P<0.01),而COS预处理能显著抑制金黄色葡萄球菌侵染所致的MUC4基因相对表达量的下调(图2-B;P<0.05),而MUC2基因相对表达量极显著下降(图2-A;P<0.01)。
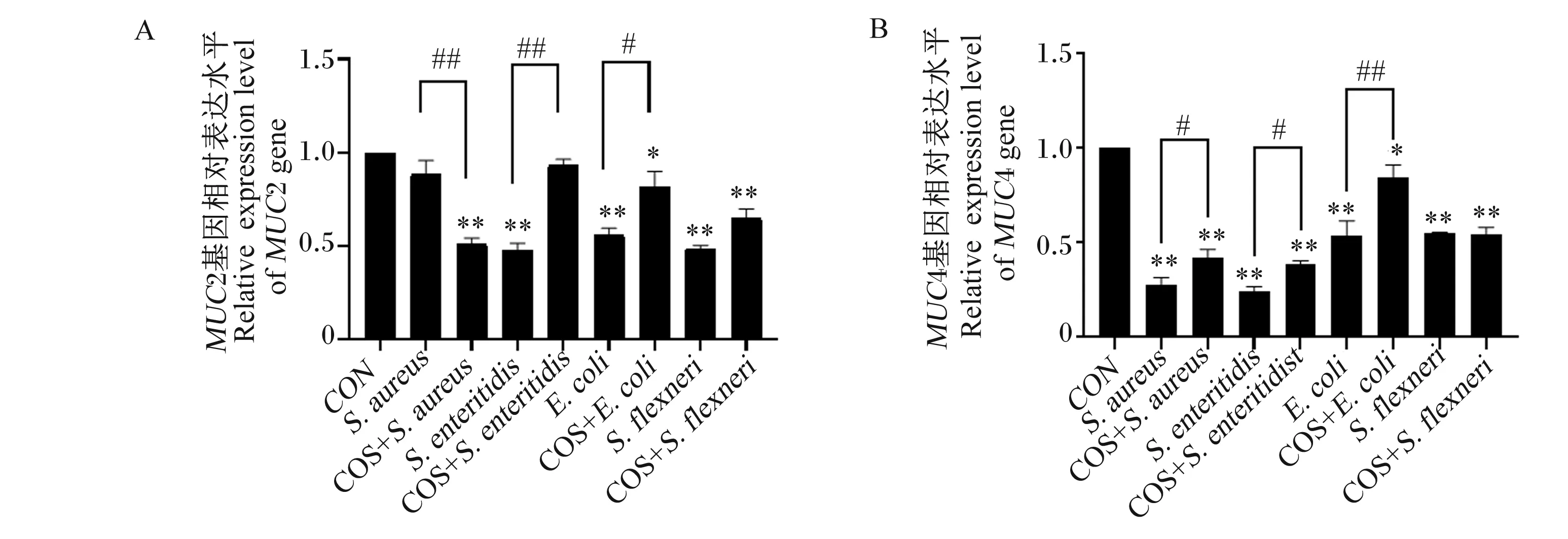
A:MUC2基因相对表达量;B:MUC4基因相对表达量。与对照组相比,** P<0.01, * P<0.05;与相应COS预处理组相比,## P<0.01, # P<0.05. 下图同。A: relative expression level of MUC2 gene; B: relative expression level of MUC4 gene. Compared with CON group: ** P<0.01, * P<0.05; compared with COS retreatment group, ## P<0.01, # P<0.05. The same as below.
2.4 COS对肠道致病菌黏附IPEC-J2细胞后紧密连接蛋白基因表达的影响
金黄色葡萄球菌、致病性大肠杆菌和福氏志贺菌与IPEC-J2细胞共培养后,细胞中Occludin基因相对表达量极显著降低(图3-A,P<0.01);而COS预处理后能显著抑制这3种细菌侵染所致的细胞Occludin基因相对表达量的下调。肠炎沙门菌侵染下细胞Occludin基因相对表达量没有显著变化(图3-A,P>0.05),但COS预处理后能使肠炎沙门菌侵染的细胞Occludin基因相对表达量显著上升(图3-A,P<0.01)。此外,只有金黄色葡萄球菌侵染能引起细胞Claudin-1基因相对表达量显著上升(图3-B;P<0.01);经COS预处理后金黄色葡萄球菌侵染的细胞Claudin-1基因相对表达量能下调至与对照组相近的水平(图3-B,P>0.05)。
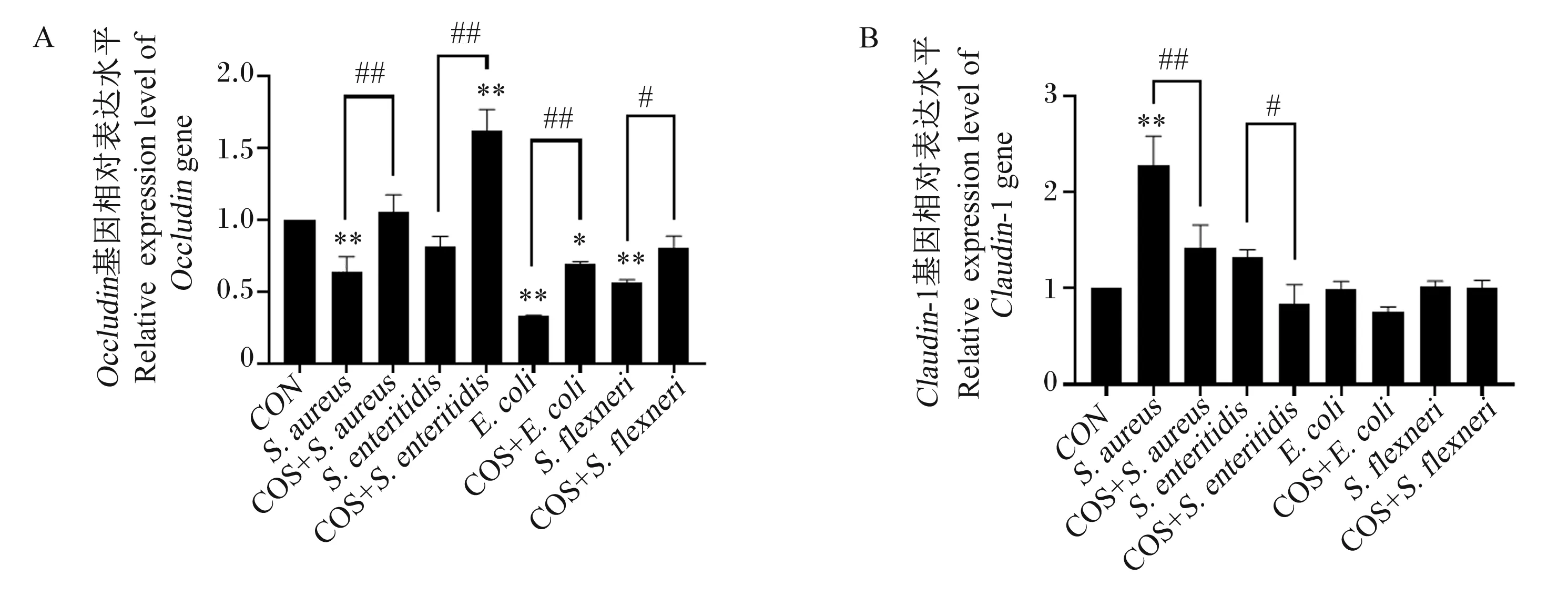
A: Occludin 基因相对表达量;B:Claudin-1 基因相对表达量。
2.5 COS对肠道致病菌黏附IPEC-J2细胞后ICAM-1和JNK基因表达的影响
金黄色葡萄球菌、致病性大肠杆菌和福氏志贺菌与IPEC-J2细胞共培养后,都能引起细胞ICAM-1基因相对表达量的极显著升高(图4-A;P<0.01);但COS预处理后能极显著抑制这4种细菌侵染所致的细胞ICAM-1基因相对表达量的上调(图4-A;P<0.01)。金黄色葡萄球菌侵染能引起细胞JNK基因相对表达量极显著上升(图4-B;P<0.01),其余3种菌侵染则无显著影响(图4-B;P>0.05);经COS预处理后能使金黄色葡萄球菌侵染的细胞JNK基因相对表达量下调至与对照组无显著差异的水平(图4-B;P>0.05);对于福氏志贺菌侵染的细胞,COS预处理组与对照组相比细胞JNK基因相对表达量显著下调(图4-B;P<0.05)。
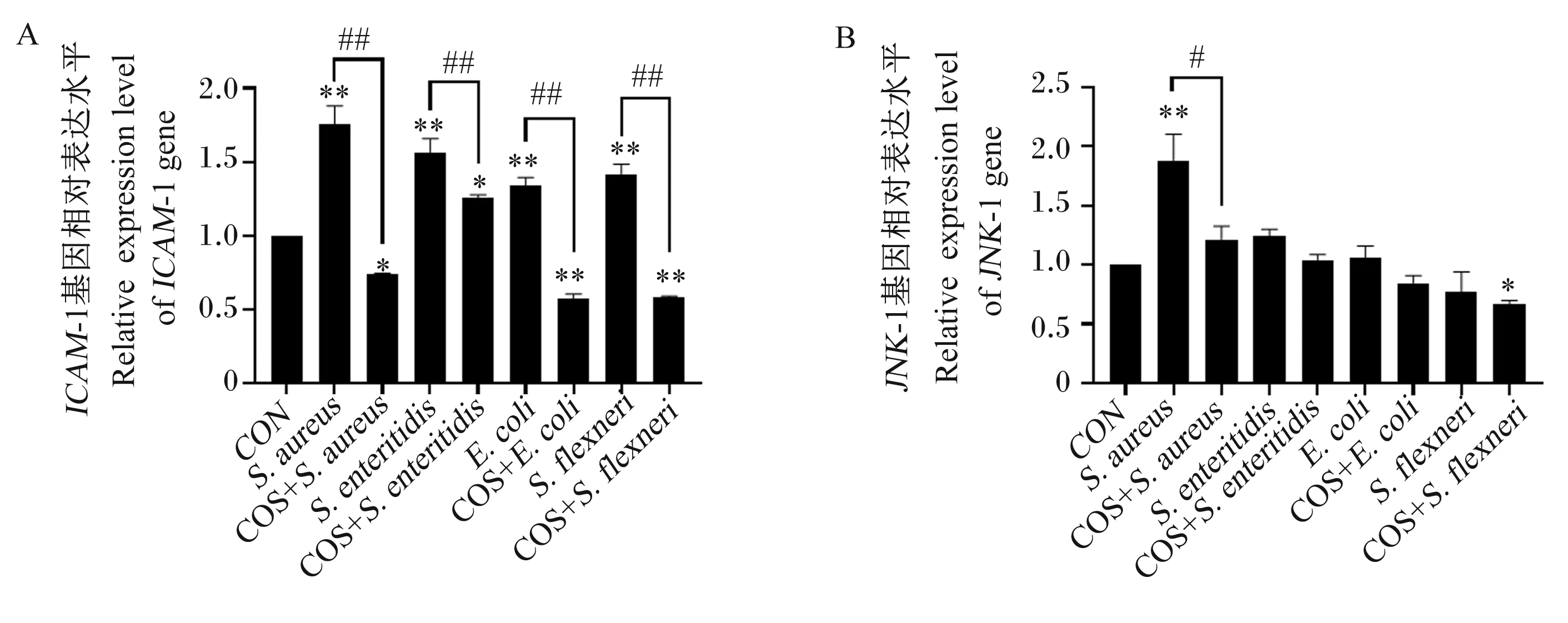
A: ICAM-1基因相对表达量;B:JNK基因相对表达量。
3 讨 论
赵倩等[14]通过共培养法证实质量浓度为100 mg/mL的COS对猪源大肠杆菌和沙门氏菌均具有抑菌作用。曹维强等[15]研究发现平均相对分子质量为609的COS对大肠杆菌和金黄色葡萄球菌的MIC为0.3%,对副伤寒甲沙门氏菌、宋内志贺菌的MIC为0.1%。张金豫[16]研究发现多种单体COS的混合物含量为16~32 mg/mL时,对大肠杆菌、金黄色葡萄球菌、枯草芽孢杆菌4种细菌均有抑制作用,但对沙门氏菌的MIC仅为6.87 mg/mL。本研究显示COS对金黄色葡萄球菌、肠炎沙门菌、福氏志贺菌均具有体外抑菌作用,MIC为64 mg/mL,MBC大于256 mg/mL;其中对致病性大肠杆菌的抑菌和杀菌效果较明显,MIC仅为16 mg/mL,MBC为128 mg/mL,说明COS对肠道致病菌具有良好的抑制作用。
COS的抑菌活性与其分子质量密切相关,小分子质量的COS可进入大肠杆菌细胞内部,与细胞内带负电的DNA结合,阻碍DNA的进一步复制,从而抑制细菌生长[17]。本研究使用的COS平均分子质量为2 000 u,可显著抑制金黄色葡萄球菌、肠炎沙门菌、致病性大肠杆菌、福氏志贺菌黏附IPEC-J2细胞,抑制率能达到34%以上。而高浓度的COS(800 μg/mL)对致病性大肠杆菌的黏附抑制率最高,可达64.63%。
MUC主要由动物肠上皮的杯状细胞合成和分泌,它一方面能够阻止微生物与上皮细胞直接接触,另一方又为微生物提供黏附位点[18-19]。分泌型黏蛋白MUC2是构成肠道黏液的最主要成分,结合型黏蛋白MUC4是一种跨膜蛋白,两者在肠道中内部厚层和外层以发挥双重作用,形成凝胶样黏液保护肠道。细菌及其代谢产物可刺激杯状细胞大量释放MUC[20],但大肠杆菌等致病菌对MUC2有快速降解作用,从而破坏黏液层的完整性[21]。沙门菌可触发干扰素-γ受体信号而影响黏液释放到肠腔,福氏志贺菌则可影响MUC的基因转录[22]。在试验性结肠炎模型中,COS处理缓解葡聚糖硫酸钠(DSS)诱导的MUC2基因表达下调,表明其能通过减少杯状细胞的破坏,提高黏液层厚度,维护肠道屏障[23]。在本试验中,肠炎沙门菌、致病性大肠杆菌和福氏志贺菌侵染后,MUC2和MUC4基因相对表达量降低,而COS预处理能显著逆转上述变化。这说明COS对肠炎沙门菌、致病性大肠杆菌作用下的肠上皮细胞具有较好的提高黏液屏障、阻止细菌直接黏附细胞进行侵害的作用。
紧密连接蛋白是上皮细胞和内皮之间最顶端的黏附蛋白,紧密连接蛋白在维持上皮屏障功能中起着重要作用。Occludin是其中的主要成分,其表达和定位对肠黏膜屏障功能的维护具有重要作用。冯洒然等[24]发现金黄色葡萄球菌能引起皮肤上皮细胞紧密连接蛋白Claudin-1、封闭蛋白-4(Claudin-4)和闭锁小带蛋白-1(ZO-1)的表达上调,但是随后出现紧密连接蛋白的表达下调及紧密连接屏障功能的破坏,这种短暂的增强是角质形成细胞的一种早期保护反应,而时间的长短与接种细菌数量也有关。Pope等[25]研究认为Claudin-1在肠上皮细胞中的过度表达能影响杯状细胞的分化和MUC2的合成。在本试验中,金黄色葡萄球菌、致病性大肠杆菌和福氏志贺菌单独处理使细胞Occludin基因相对表达量显著下降,而COS预处理后,Occludin基因相对表达量恢复到与对照组相近的水平。由此可见,COS能较好地缓解这3种细菌黏附肠上皮细胞造成的屏障损伤。
细菌及其产物可刺激上皮细胞释放黏附分子,如脂多糖可诱导细胞黏附和ICAM-l表达,以维持肠道完整性和调节免疫细胞向肠道的迁移,释放炎性介质[26]。陈爽[23]发现COS能抑制DSS诱导的小鼠结肠黏附分子ICAM-1和血管细胞黏附分子1(vzascular cell adhesion molecule-1,VCAM-1) mRNA的表达水平。本研究发现,4种肠道致病菌都引起细胞ICAM-1基因相对表达量的升高;在COS预处理后同样能极显著降低ICAM-1基因相对表达量,抑制细菌黏附造成的炎性反应,保护肠道黏膜功能。
Suzuki等[27]和Kojima等[28]证明了ICAM-1基因表达受到JNK的正调控。JNK的激活也可以负调节MUC2转录[29]。Deng等[30]研究发现,抑制JNK的活化可以保护紧密连接不被DSS诱导破坏,减轻上皮屏障功能障碍。程晓鸥[31]发现金黄色葡萄球菌入侵可引起JNK入核,核内磷酸化c-Jun增多。张坤[32]研究表明,JNK对大肠杆菌侵染细胞能够激活细胞内的转录因子,从而引起免疫调控相关基因的表达。本试验中只有金黄色葡萄球菌能引起细胞JNK基因相对表达量极显著上升,COS预处理后对金黄色葡萄球菌共培养的细胞JNK基因相对表达量下调。由此可见,COS对4种细菌作用下细胞ICAM-1基因相对表达量的升高具有恢复作用,其中COS对金黄色葡萄球菌侵染后细胞ICAM-1基因相对表达量的下调的缓解作用可能通过JNK通路介导。
4 结 论
COS对致病性大肠杆菌的抑菌和杀菌效果较明显,MIC为1.6 mg/mL;对金黄色葡萄球菌、肠炎沙门菌、福氏志贺菌也具有抑菌作用,MIC为6.4 mg/mL。COS对这4种致病菌均有良好抑菌和黏附抑制作用,其中对致病性大肠杆菌的杀菌效果明显。COS能显著抑制致病性大肠杆菌对IPEC-J2细胞的黏附,机制与ICAM-1基因表达抑制、MUC和紧密连接蛋白基因表达上调有关。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
九江学院学报(自然科学版)(2022年2期)2022-07-02
健康体检与管理(2022年2期)2022-04-15
家庭医药·快乐养生(2020年11期)2020-12-03
大众科学·下旬(2019年6期)2019-09-10
科研成果与传播(2019年3期)2019-09-10
江苏农业科学(2015年11期)2016-01-27
中国动物保健(2015年4期)2015-10-21
湖北畜牧兽医(2014年7期)2014-09-24
