
大气CO2 浓度缓增对稻田土壤甲烷氧化过程的影响*
2022-06-09 01:17:00沈李东田茂辉杨王挺金靖昊王昊宇胡正华
土壤学报 2022年2期
刘 心,沈李东,田茂辉,杨王挺,金靖昊,王昊宇,胡正华
(南京信息工程大学生态系,南京 210044)
甲烷(CH)是仅次于二氧化碳(CO)的长寿命温室气体,对全球温室效应的贡献率约为17%。甲烷在100 年尺度的全球增温潜势是等质量CO的28 倍。自工业革命以来,大气中甲烷浓度增加了150%,其排放量仍以每年1.0%的速度在递增。稻田是重要的甲烷排放源之一,其每年甲烷排放量约占全球排放总量的10%~20%。微生物介导的甲烷好氧氧化被认为是控制稻田甲烷排放的重要途径,可氧化产甲烷菌产生的40%~90%的甲烷。
根据细胞结构和功能及系统进化发育关系,甲烷氧化菌可分为三大类。第一类包括Ⅰ型和Ⅹ型甲烷氧化菌,属于变形菌纲,主要有甲基杆菌()、甲基球菌(、甲基单胞菌()、甲基微菌()和甲基八叠球菌()等10 个属。第二类为II 型甲烷氧化菌,属于变形菌纲,主要包括甲基弯曲菌()和甲基包囊菌()等5个属。 第三类甲烷氧化菌属于疣微菌门(Verrucomicrobia),主要由嗜热嗜酸菌组成。有研究发现,稻田土壤中甲烷氧化菌的主要类型是II 型菌。但也有研究表明,I 型菌主要分布在水稻的根际和水土交界面的表层土壤中,而II 型菌则更适应低氧环境,其在下层缺氧区占优势地位。
CO作为最重要的温室气体,对气候变暖的贡献最大。气候模型预测表明,至21 世纪末大气中CO浓度可能会翻一番。大气CO浓度升高会极大地影响陆地生态系统的性质和功能。在水稻不同生长期,土壤环境因子会发生改变,并与大气CO协同作用后对土壤甲烷氧化过程产生影响。土壤中的CO浓度是大气中的10 倍~15 倍,因此大气CO浓度升高几百μL·L很难直接改变土壤微生物群落结构。但有证据表明,高浓度的CO能够促进植物生长发育,并通过增加植物向土壤的碳输入以及改变其他相关土壤理化性质(如氮素水平、氧气含量)等影响土壤微生物群落结构和活性。可见,大气CO浓度升高可能会对稻田甲烷氧化菌的活性、丰度和群落结构产生间接影响,进而改变稻田甲烷排放量。有研究表明,较高的大气CO浓度会使土壤中甲烷的氧化量减少,进一步加剧气候变化。然而也有研究发现,大气CO浓度升高能够促进稻田土壤甲烷氧化菌的生长。因此,当前关于大气CO浓度升高对稻田甲烷氧化过程的影响结论仍存在争议,其影响机理尚不明确。
大气CO浓度是长时间缓慢增加的过程,而不是短期内增至某一浓度后固定不变。目前尚少见CO浓度缓增对土壤甲烷氧化菌影响的相关研究。本研究在背景CO浓度基础上设置了CO浓度缓增处理,更加真实地模拟大气CO浓度升高的情境。此外,高通量测序方法(如Illumina MiSeq)在研究土壤微生物群落中更具有优势,能够对微生物群落特征提供更全面的解释。然而该方法较少应用于研究稻田土壤甲烷氧化菌群落的动态变化。本研究采用Illumina MiSeq 测序技术以及定量PCR 等手段,从基因、群落、活性等多个层次上解析CO浓度缓增对稻田土壤甲烷好氧氧化过程的影响及其作用机理。
1 材料与方法
1.1 试验区概况
试验区设置在江苏省农业气象重点实验室的稻田试验地(32.16 °N,118.86 °E),属亚热带湿润气候区,年平均降水量1 100 mm,多年平均气温15.6 ℃。供试土壤为潴育型水稻土,灰马肝土属,耕层土壤质地为壤质黏土。供试水稻品种为南粳9108,全生育期为149~153 d。稻田土壤的氮肥年施用量为250 kg·hm。水分管理方式为生育前期淹水、中期烤田、后期干湿交替。
1.2 土壤样品采集与分析
本研究依托于开顶气室(Open Top Chambers,OTC)组成的CO浓度升高自动调控平台,该平台于2016 年建成并运行,包括OTC、CO传感器、自动控制系统及通气装置。每个OTC 内配置CO传感器(GMM222 传感器,芬兰)及温湿度自动记录仪。通过计算机程序实现OTC内CO浓度的自动监测和调节,以确保OTC 内CO浓度维持在目标浓度值。CO气源为高纯CO钢瓶。
以大气CO浓度为对照(Ambient CO,AC),设置CO浓度缓增处理(Elevated CO,EC),每个处理各3 个重复。EC 处理为在背景CO浓度基础上,自2016 年起每年增加40 μL·L。于2019 年采集水稻4 个关键生育期即:分蘖期、拔节期、扬花期和乳熟期0~5 cm 的根际土壤。该年EC 处理的CO浓度为AC+160 μL·L。采用烘干法测定土壤含水率;以玻璃电极法测定土壤pH;以比色法测定土壤无机氮( NH-N、NO-N和NO-N);土壤有机质含量的测定方法参考《土壤农化分析》。
1.3 甲烷氧化潜势测定
取约3 g 新鲜土样置于12 mL 玻璃瓶(Labco,英国)中,并向瓶内加入5 mL 去离子水后,用橡胶塞密封。随后,用纯甲烷进行等体积置换,使甲烷含量占比约10%(体积比)。将玻璃瓶置于摇床以35 ℃,120 r·min振荡培养。期间,每隔12 h 用50% ZnCl溶液终止一批反应,直至最后一批样品于第72 h 培养结束。用气相色谱仪(Agilent 7890B,美国)测定玻璃瓶中顶空甲烷浓度。最后,根据甲烷浓度随时间减少进行线性拟合,计算甲烷氧化潜势。
1.4 DNA 提取和PCR 扩增
根据 Power Soil DNA Isolation kit 试剂盒(MoBio,美国)说明书对不同处理下稻田土壤样品的总DNA 进行抽提。DNA 浓度和纯度利用超微量分光光度计(NanoDrop2000,美国)进行测定,并利用1%琼脂糖凝胶电泳检测DNA 提取质量。利用特 异 性 引 物 A189F (5′-GGNGACTGGGACTTC TGG-3′)和mb661R(5′-CCGGMGCAACGTCYTTAC C-3′)对甲烷氧化菌基因进行PCR 扩增。扩增程序如下:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,30 个循环;最后以72 ℃延伸10 min。PCR 反应体系为20 μL:4 μL 5×astPfu 缓冲液,2 μL 2.5 mmol·LdNTPs,0.8 μL正 向 引 物( 5 μmol·L), 0.8 μL 反 向 引 物(5 μmol·L),0.4 μL FastPfu 聚合酶,0.2 μL BSA(牛血清蛋白),以及10 ng DNA 模板。
1.5 Illumina Miseq 测序
利用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences,USA)进行 PCR 产物纯化;利用QuantiFluor™-ST(Promega,USA)进行产物检测定量。根据Illumina MiSeq 平台(Illumina,USA)标准操作规程将纯化后的扩增片段构建PE 2×300的文库(上海美吉生物医药科技有限公司)。
当前,数字管理已深入人心。大数据的价值不仅在于其原始价值,更在于数据的连接、扩展、再利用和重组,如何将大数据的相关性特征更好地应用于房地产估价行业和机构管理,离不开智慧管理。从行业来看,行政管理部门和行业协会可通过房地产估价大数据监管模型、信用信息共享交换平台、网络新闻评论数据库等智慧管理手段实现行业监管。从机构来看,在市场数据库、估价实务知识库、估价基础知识库和网络管理大平台的数字化建设基础上,还要对高级管理者特殊才能、高级技能人员经验、组织惯例与本能、产业链适应性资源、产品品牌价值等智慧资源进行系统管理,以实现机构的可持续发展[7—8]。
1.6 序列分析
原始序列使用 Trimmomatic 软件质控,以FLASH 软件进行拼接,具体步骤如下:(1)设置50 bp 的窗口,若窗口内的平均质量值低于20,从窗口前端位置截去该碱基后端所有序列,之后再去除质控后长度低于50 bp 的序列;(2)根据重叠碱基overlap 将两端序列进行拼接(最大错配率为0.2,长度需大于 10 bp);(3)根据序列首尾两端的barcode 和引物将序列拆分至每个样本并精确匹配。使 用 UPARSE 软 件( version 7.1 http ://drive5.com/uparse/)以 86%的相似度对序列进行OTU 聚类,同时去除单序列和嵌合体。最终通过对比数据库,利用 RDP classifier(http://rdp.cme.msu.edu/)对每条序列进行物种分类注释。甲烷氧化菌的基因序列已上传至美国国家生物技术信息中心(National Center of Biotechnology Information,NCBI),序列登录号为PRJNA612131。
1.7 定量PCR
使用特异性引物A189F 和mb661R,通过定量PCR(quantitative PCR,qPCR)测定甲烷氧化菌基因的丰度(PCR 反应体系和反应条件参见1.4 部分)。利用质粒标准样品稀释7 个浓度梯度,绘制标准曲线。根据标准曲线得到每个样品的基因拷贝数(每个样品3 个重复)。
1.8 数据处理
采用美吉生物云平台对测序数据进行分析。使用SPSS 25.0 软件的检验分析不同处理间各指标(甲烷氧化活性、甲烷氧化菌数量、多样性等)的差异及其显著性。使用SPSS 25.0 软件对各理化指标和微生物指标之间的关系进行 Pearson 相关性分析。
2 结 果
2.1 大气CO2 浓度缓增对稻田土壤理化因子的影响
不同CO浓度处理下各生育期稻田土壤的理化性质见表1。结果显示,在不同CO浓度处理下,四个关键生育期的土壤含水率和pH 在较小的范围内波动。在分蘖期,AC 和EC 处理下土壤中DOC含量未见明显变化,但EC 处理下土壤中 NH-N、NO-N和NO-N含量均显著低于AC 处理。在拔节期,EC 处理的NO-N含量显著高于(< 0.05)AC处理,而其他土壤理化因子未见有显著变化。在扬花期,与AC 相比,EC 处理的土壤有机碳与DOC含量显著提高(< 0.05),但 NH-N 和NO-N含量显著降低(< 0.05)。在乳熟期,EC 处理下土壤中DOC 含量较AC 有所增加,但未达到显著性水平。
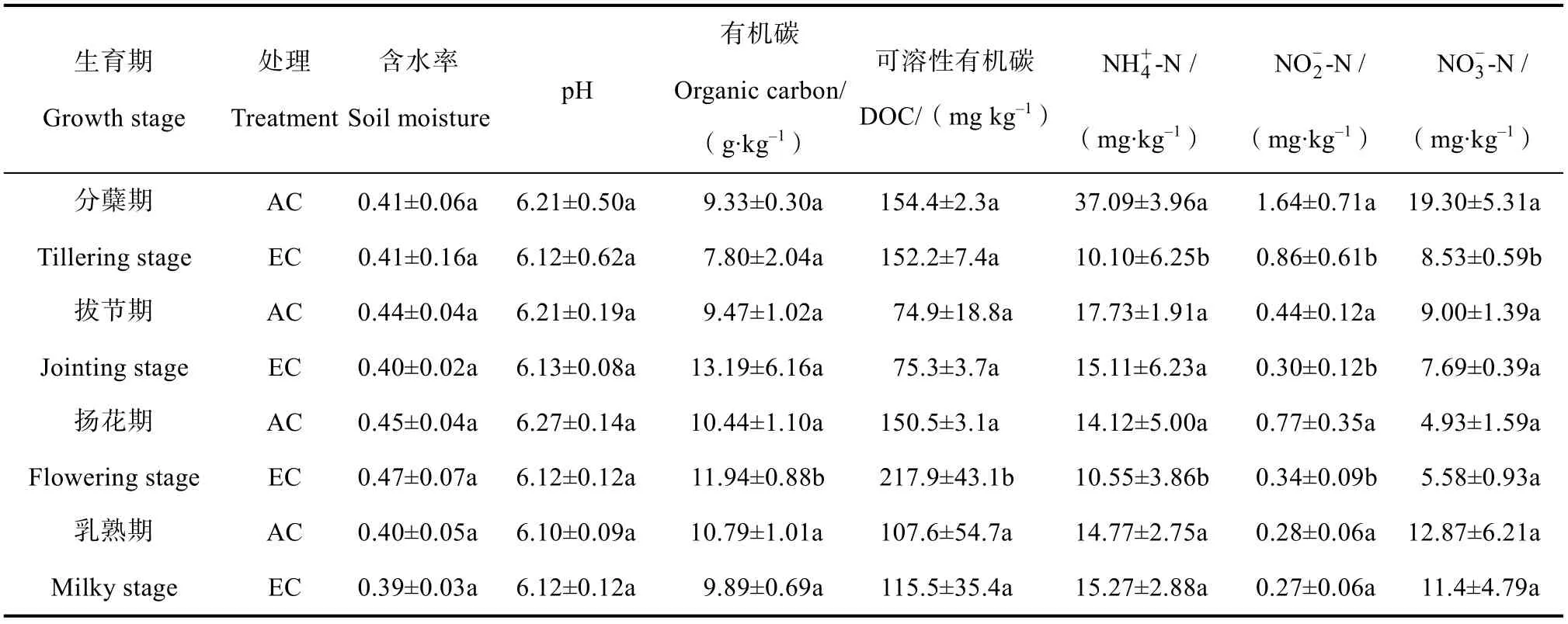
表1 不同CO2 浓度处理下各生育期稻田土壤的理化性质Table 1 Physiochemical properties of paddy soils during different rice growth stages under different CO2 treatments
2.2 大气CO2 浓度缓增对稻田土壤甲烷氧化潜势及甲烷氧化菌丰度的影响
土壤泥浆培养试验结果显示,该稻田土壤的甲烷氧化潜势在分蘖期达到最高值,并随着水稻的生长,甲烷氧化潜势呈下降趋势(图1a)。除分蘖期外,EC 处理促进了土壤甲烷氧化潜势,且在拔节期和乳熟期达到显著性水平(< 0.05)。分蘖期的土壤甲烷氧化潜势随CO浓度的升高显著降低(<0.05)。综合每个生育期的数据,EC 处理使得稻田整体甲烷氧化潜势增加11.7%。
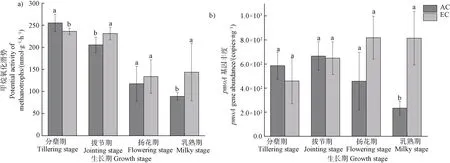
图1 不同CO2 浓度处理下各生育期稻田土壤的甲烷氧化潜势(a)及甲烷氧化菌pmoA 基因丰度(b)Fig. 1 Potential activity (a) and pmoA gene abundance (b) of methanotrophs in paddy soils during different rice growth stages under different CO2 treatments
2.3 大气CO2 浓度缓增对稻田土壤甲烷氧化菌pmoA 基因多样性的影响
利用OTU 水平上的Sobs 指数、香农指数和Chao 1 指数,评估不同CO浓度处理下土壤中甲烷氧化菌丰富度和多样性的变化。数据分析结果显示,单个样品中OTU 数介于40~189 之间。所有样品OTU 的稀释曲线均接近饱和状态,表明本研究的测序深度足以描述甲烷氧化菌基因的多样性。研究发现,各时期的甲烷氧化菌基因多样性随大气CO浓度升高呈降低趋势。其中在乳熟期,EC 处理下土壤中甲烷氧化菌的多样性显著低于(< 0.05)AC 处理。但在其他三个生育期,不同处理下土壤中基因多样性差异不显著(图2)。
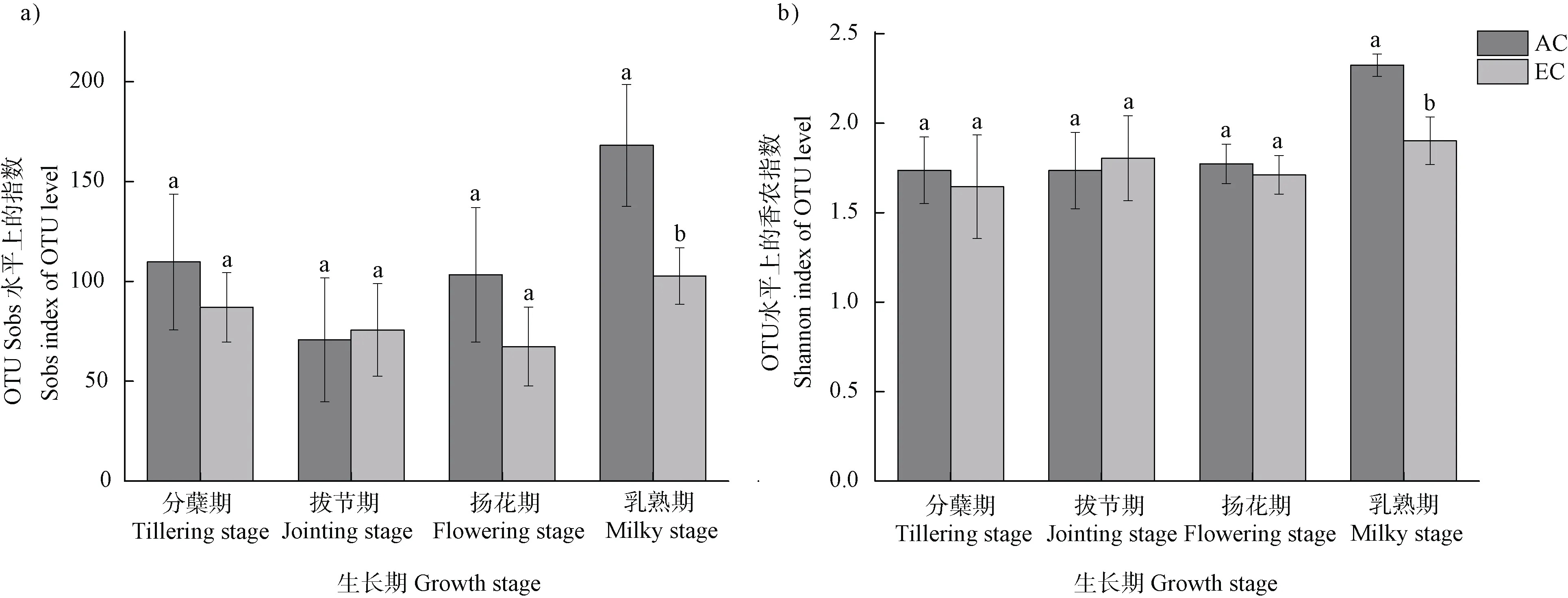
图2 不同CO2 浓度处理下各生育期稻田土壤中pmoA 基因的Sobs 指数(a)和香农指数(b)Fig. 2 The Sobs index(a)and Shannon index(b)of pmoA genes in paddy soils during different rice growth stages under different CO2 treatments
2.4 大气CO2 浓度缓增对稻田土壤甲烷氧化菌群落结构的影响
以属为水平对不同CO浓度处理下稻田土壤中甲烷氧化菌的群落组成进行分析。结果发现已知菌属主要包括Ⅰ型的甲基八叠球菌()、甲 基 单 胞 菌()、 甲 基 球 菌(、甲基杆菌()和Ⅱ型的甲基包囊菌()(图3)。在AC 处理下,土壤中Ⅰ型甲烷氧化菌占比37.23%,Ⅱ型菌占比39.95%;在EC 处理下,Ⅰ型占比50.2%,高于占比30.43%的Ⅱ型菌。因此,大气CO浓度升高改变了甲烷氧化菌的优势菌群,使得Ⅱ型菌占比明显提高。甲基包囊菌属、甲基八叠球菌属和甲基单胞菌属始终是该稻田土壤中的优势菌属。在AC 处理下,土壤中这三类优势甲烷氧化菌属的相对丰度分别为39.95%、17.86%和8.69%;而在EC 处理下,这三种菌属的相对丰度则分别为 30.43%、20.62%和18.77%。此外,土壤中还存在甲基球菌和甲基杆菌,以及其他未被分类的菌属,但其相对丰度较低(图3)。

图3 不同CO2 浓度处理下各生育期稻田土壤甲烷氧化菌的群落组成Fig. 3 Community composition of methanotrophs in paddy soils during different rice growth stages under different CO2 treatments
对不同CO处理下土壤中甲烷氧化菌群落组成进行组间差异显著性检验(图4 a),发现某些环境序列和未被分类的菌属的相对丰度存在显著性差异(< 0.05)。但在乳熟期,EC 处理下使得甲基单胞菌的相对丰度显著高于(< 0.05)AC 处理,而甲基包囊菌的相对丰度则显著低于(< 0.05)AC 处理(图4b)。总体而言,甲烷氧化菌群落在两种CO浓度处理间,存在显著差异(图5,= 0.039)。
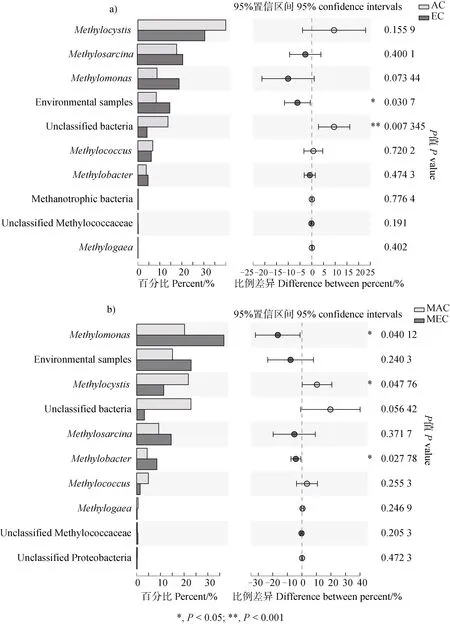
图4 不同CO2浓度处理下稻田土壤中甲烷氧化菌在属水平上的总体群落结构差异(a)以及在和乳熟期的群落结构差异(b)Fig. 4 Variation of methanotrophic community composition at genus level in paddy soils during rice growth period(a)and at milky stage(b)under different CO2 treatments
2.5 影响稻田甲烷氧化菌群落及其活性的主要环境因子
RDA 结果显示,土壤pH 和NO-N 含量对甲烷氧化菌的群落结构有显著影响(< 0.05)(图5)。不同CO浓度处理下土壤的甲烷氧化潜势和基因丰度与DOC 含量、NH-N 含量呈正相关,而甲烷氧化潜势又与pH 呈负相关,但均未达到显著性水平(表2)。本研究未发现单一的环境因子对甲烷氧化潜势和基因丰度、多样性的变化有显著影响。此外,甲烷氧化潜势和基因多样性存在显著正相关(< 0.05)。
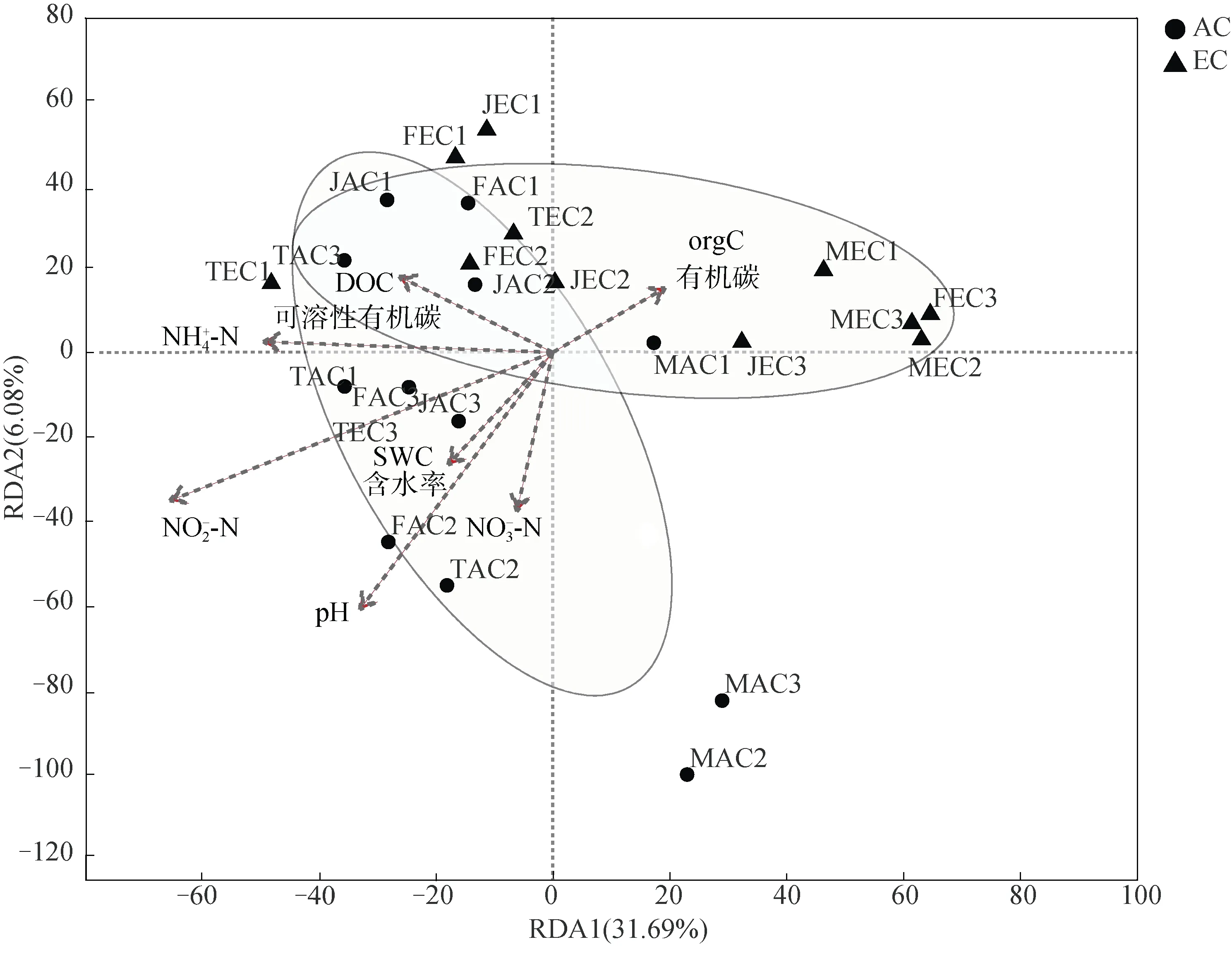
图5 不同CO2 浓度处理下稻田土壤中甲烷氧化菌群落结构与环境因子关系的RDA 图Fig. 5 Redundancy analysis(RDA)ordination plots showing the relationship between the community structures of methanotrophs and environmental factors in paddy soils under different CO2 treatments
3 讨 论
甲烷是甲烷氧化菌唯一的碳源和能源。据报道,土壤中甲烷浓度是影响稻田甲烷氧化潜势和甲烷氧化菌群落的重要因素。大量研究表明,土壤中甲烷氧化速率随甲烷浓度的升高而增加。大气CO浓度升高会促进稻田中甲烷的产生量。这主要是由于CO浓度升高刺激了水稻的生长发育,增加了其根系生物量和分泌物,导致向土壤中释放更多的含碳物质。这给产甲烷菌提供了更多的底物来源。本研究发现,大气CO浓度缓增增加了某些生长期稻田根际土壤甲烷氧化菌的丰度以及甲烷氧化潜势(图1)。相关性分析显示,甲烷氧化潜势和基因丰度均与DOC 含量呈正相关关系(表2)。这说明大气CO浓度升高间接导致土壤DOC 含量增加,使产甲烷潜势增强,进而给甲烷氧化菌提供了更多底物甲烷来源。土壤中相对较高浓度的甲烷有利于甲烷氧化潜势和甲烷氧化菌丰度的增加。
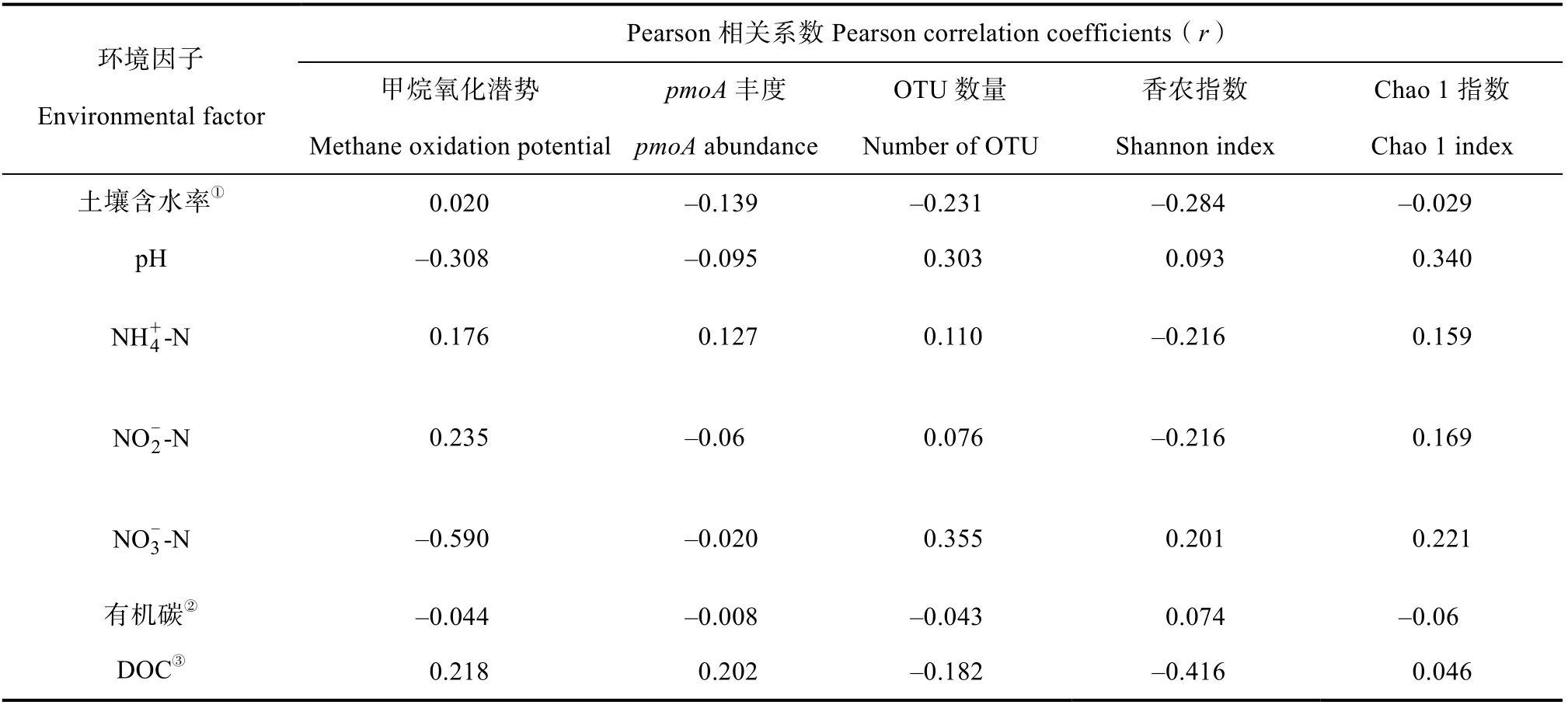
表2 不同CO2 浓度处理下稻田土壤的甲烷氧化潜势及pmoA 基因丰度、多样性与环境因子的相关性Table 2 Potential correlations between the potential activity of methanotrophs,pmoA gene diversity and abundance and different environmental factors in paddy soils under different CO2 treatments
与此同时,氧气的可获得性是甲烷氧化的重要限制因子。大气CO浓度升高可使水稻根系的通气组织愈加发达,继而增加大气向水稻根际O的输送。这在一定程度上可以提高水稻根际的甲烷氧化潜势。早期,王义琴等利用微根窗技术对植物进行观测,发现在大气CO浓度倍增条件下,植物的根长和根数量与自然环境生长的植物相比,分别增加了16%和34%。本研究结果发现,在扬花期和乳熟期的土壤中,甲烷氧化菌丰度与同时期的甲烷氧化潜势呈相同的变化趋势:均随大气CO浓度的升高而显著增加(< 0.05)(图1)。该结果很可能是由于在水稻生长的中后期,大气CO浓度升高下水稻根系生长更加发达,使得根际区域O含量增加,从而刺激了甲烷氧化菌的生长与活性。先前的相关研究也发现,大气CO浓度升高会促进稻田土壤甲烷氧化菌活性及其基因丰度的增加。然而也有研究发现,大气CO浓度升高会增强叶片的水分利用效率,使得土壤水分增加,进而通过影响土壤中O扩散速率抑制甲烷氧化活性。但这种情况多发生于森林、草地等生态系统中。稻田土壤长时间处于淹水条件,水分较为充足。大气CO浓度升高所引起的土壤含水量的微小增加,可能不足以改变稻田中的气体扩散速率。本研究中稻田表层土壤的含水率较高,且并未随CO浓度升高发生明显改变(表1)。
氮素含量是影响稻田甲烷氧化的另一重要环境因子。本研究发现,大气CO浓度升高导致了土壤中 NH-N 和NO-N含量的减少(表1)。有研究表明,通过长达15 年的CO富集试验,大气CO浓度升高会造成稻田生态系统中 NH-N 的大量损失,这可能是由于 NH的厌氧氧化和铁的还原增强所致。土壤中 NH-N 的减少还可能与作物根系生长对铵态氮的吸收增强有关。同时,大气CO浓度升高会降低土壤NO-N的积累,这是因为稻田根系生物量和分泌物的增加促进了反硝化活性的增强。稻田土壤中较高浓度的 NH会通过增加硝化作用来抑制甲烷氧化菌的生长及活性。Adamsen 和King在土壤培养实验中发现,较高浓度的NO-N也会抑制土壤甲烷氧化活性。但其抑制作用的机理尚不明确,可能是由于NO本身或通过NO还原产生NO对甲烷氧化菌的毒害作用。通常情况下,稻田土壤的本底 NH-N 含量较高(氮肥施用所致)。为此,本研究中大气CO浓度升高所致土壤 NH-N 和NO-N积累的减少,可能有助于缓解氮素对稻田甲烷氧化的抑制作用。
随着大气CO浓度的升高,甲烷氧化菌的优势菌群发生了改变:从Ⅱ型菌(相对丰度为39.95%)转变为Ⅰ型菌属(相对丰度为50.2%)。因此,大气CO浓度升高促使了土壤中Ⅱ型菌向Ⅰ型菌的演替。AC处理的优势菌属为甲基包囊菌(Ⅱ型)、甲基八叠球菌和甲基单胞菌(均为Ⅰ型)(图3)。这与先前稻田土壤中甲烷氧化菌的主要类型是Ⅱ型菌的结果一致。据报道,甲基弯曲菌和甲基包囊菌(均为Ⅱ型)的相对丰度会随大气CO浓度升高而降低。研究表明,在高 NH条件下,甲基孢囊菌(Ⅱ型)的竞争力高于Ⅰ型菌。大气CO浓度升高所导致NH-N 含量的减少,很可能削弱了甲基孢囊菌的竞争力,使其丰度下降(图4 a)。除氮素含量外,稻田土壤中的O浓度和营养条件的变化亦可能会改变甲烷氧化菌的群落结构。以往的研究表明,Ⅰ型菌在高氧和营养充足的环境中有较强的竞争优势,而Ⅱ型菌更适应在低氧和寡营养的环境下生长。如前所述,大气CO浓度升高会促进水稻根系分泌物和通气组织容积的增加,这为Ⅰ型菌的生长提供了更为合适的生存环境。
4 结 论
大气CO浓度缓增促进了稻田土壤甲烷氧化潜势的增加,以及甲烷氧化菌的生长。同时,大气CO浓度升高显著改变了土壤中甲烷氧化菌的群落结构,使得优势菌属由起初的Ⅱ型菌转变为Ⅰ型菌。大气CO浓度升高会促进作物的生长,导致稻田土壤中甲烷浓度和根际环境通气性的增加,并调节土壤中的氮素水平。以上多个环境因子的综合作用很可能是稻田甲烷氧化潜势、甲烷氧化菌丰度与群落结构发生改变的重要原因。
猜你喜欢
农业灾害研究(2022年1期)2022-05-07 01:31:04
选煤技术(2022年1期)2022-04-19 11:15:00
趣味(作文与阅读)(2021年5期)2021-08-19 02:48:54
能源工程(2021年2期)2021-07-21 08:39:54
军民两用技术与产品(2021年10期)2021-03-16 06:05:08
今日农业(2020年21期)2020-12-19 13:52:28
水上消防(2020年1期)2020-07-24 09:26:02
作文大王·低年级(2019年2期)2019-01-23 11:35:18
创新作文(小学版)(2018年19期)2018-11-30 01:56:08
疯狂英语·新读写(2018年3期)2018-11-29 22:37:11
