
干旱对碧玉杨幼苗水氮利用与同化物分配的影响
2022-06-08 07:12:38尚佳州赵瑜琦王卫锋高钿惠宗毓铮
干旱区研究 2022年3期
尚佳州, 赵瑜琦, 王卫锋, 高钿惠, 宗毓铮
(1.山西农业大学林学院,山西晋中 030801;2.山西农业大学农学院,山西晋中 030801)
杨树(Populusspp.)是我国北方地区的重要树种,因其速生、造林成活率高等特性,20世纪50年代以来,一直是“三北”地区生态防护林的主要树种,该地区以小叶杨(P.simonii)、群众杨(P.×popularis‘35-44’)等树种营造的人工乔木林,可能是当地土壤干层形成的重要原因,而土壤干层又抑制了地上杨树人工林植被生长,并形成了大面积“小老树”退化林分[1-3]。未来气候变化下该地区的干旱频率与强度可能会增加[4],将进一步限制杨树人工林生产力甚至导致林分死亡。
干旱下杨树的生理生态响应过程已有不少研究,土壤水分有效性降低可导致杨树叶片气孔关闭、光合速率降低,并使得个体生长受到抑制[5],体内有限的碳氮资源分配模式发生改变[6],来维持整株水分平衡以及个体存活[7-8],这些生理生态过程的响应程度与树种、器官功能、生长阶段等因素有关[9]。多数杨树树种对土壤水分胁迫较为敏感,可调节根系空间分布以促进吸水[10]、木质部导管直径减小维持水分运输安全性[11]、以及减小叶片数和叶面积降低水分散失,而抗旱性较强的杨树则可以通过灵敏的气孔开闭提高水分利用效率[12-13]。干旱下群众杨叶片光合作用显著下降且茎干物质分配略有增加[14],而干旱下小叶杨的根冠比和叶茎根中可溶性糖含量均显著增加,且在复水后出现明显的光合作用补偿效应[15]。虽然,干旱地区树木茎中可溶性糖含量增强的形成机理还存在争议[16],但该现象参与的渗透调节通常被认为有助于修复导管栓塞进而维持水分长距离运输等生理过程[17]。然而,干旱下杨树的生理生态响应过程还不够清楚,种间抗旱策略差异也需要进一步明确。
据报道欧美杨新品种碧玉杨(P. ×euramericana‘Biyu’)抗寒抗旱性强,可在内蒙古中西部[18]、甘肃山丹县[19]等干旱地区良好生长。田间观察发现充分灌水下碧玉杨速生特性明显,但控制干旱下碧玉杨生长量大幅降低[20];且其根冠关系、非结构性碳积累等方面对干旱的生理生态响应仍不明确。本试验以碧玉杨扦插苗为材料,通过盆栽控制试验,研究了连续2 a土壤干旱下碧玉杨叶片光合特性、水氮利用和器官间同化物分配的响应变化,研究结果可有助于进一步明确干旱下杨树的响应机理,并为干旱半干旱区杨树人工林高效培育提供理论依据。
1 材料与方法
1.1 试验地概况
试验在山西省晋中市太谷县山西农业大学林学院实验苗圃高透光遮雨棚下进行。该苗圃位于112.57°E,37.43°N,海拔796 m;属暖温带大陆性气候,年均日照时数2527.5 h,年均气温10.4 ℃,年均降水量397.1 mm,年平均蒸发量1649 mm,无霜期176 d。供试土壤取自苗圃表层土,质地为壤土,基本理化性质为:有机质1.79 mg·g-1,全氮2.4 mg·g-1,全磷1.5 mg·g-1,pH 7.22,田间持水量为26.94%。
1.2 试验材料与处理
于2016年4月初选用高35 cm、直径30 cm的塑料桶,桶底铺1层鹅卵石及2层报纸与土壤隔离,每桶装风干土18 kg;通过插到桶底的硬质塑料管浇水,有利于土壤自下而上均匀吸水且避免土壤板结。选取生长良好粗细均匀的一年生碧玉杨苗干的中间部位剪成长约12 cm 插条,每桶扦插1 株,共60株,进行正常盆栽管理。2016年6月底开始用称重法进行持续2 a的土壤干旱处理,设置3个水分处理:(1)正常供水(CK:土壤含水量为田间持水量的75%±5%);(2)中度干旱(MS:土壤含水量为田间持水量的50%±5%);(3)重度干旱(SS:土壤含水量为田间持水量的25%±5%),每个处理20 株重复。每天傍晚进行称重补水,使各处理维持在设定的土壤相对含水量范围内,并准确记录补水量。分别于2016年9月初与2017年9月初对上述试验材料进行叶片气体交换、碳氮含量等指标测定。
1.3 气体交换参数测定
每处理随机选取6 株,于10:00—11:30 用便携式光合速率测定仪(Li-6400XT,Li-COR,USA)测定每株功能叶中部的净光合速率(Pn,μmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1)等气体交换参数。红蓝光源光强设定为1500 μmol·m-2·s-1,流速为500 μmol·s-1,叶温25 ℃,相对湿度60%~70%,外界CO2浓度(Ca)为410~430 mmol×mol-1。并计算气孔限制值(Ls)和瞬时水分利用效率(WUE)[21]。
1.4 叶片碳氮元素含量测定
选取与光合参数所测叶片相近的叶片,105 ℃杀青1 h 后75 ℃烘干至恒重,磨碎过200 目筛待用。测定时称取0.1 g 样品,用硫酸加速剂混合消煮,用凯氏定氮仪(FoodALYTD5000,OMNILab,Germany)测定叶氮含量。并计算单位面积叶氮含量、总叶氮、整株氮利用效率(NUE)和光合氮利用效率(PNUE)[21]。叶碳含量采用重铬酸钾-硫酸氧化法进行测定。
1.5 生长指标和非结构性碳含量测定
分别用游标卡尺和卷尺测定地径和株高。选取与光合参数所测叶片相近的叶片,使用扫描仪(DS50000,EPSON,Japan)测得单叶面积。收获时小心将根系冲洗干净,并整株分成根、茎、叶3部分,分别置于牛皮纸袋中,放入烘箱中杀青后75 ℃烘干至恒重,用分析天平称重并计算比叶重等参数。处理第2 年的根茎叶干样粉碎后过100 目筛,然后用苯酚-硫酸法分别测定其可溶性糖和淀粉的含量[22]。
1.6 数据分析
采用Excel 2013 进行原始数据进行初步处理;在SPSS 17.0 中用one-way ANOVA 进行单因素方差分析,及Duncan’s 新复极差法进行多重比较(显著水平α=0.05)。
2 结果与分析
2.1 干旱对碧玉杨气体交换参数的影响
2016 年与2017 年碧玉杨气体交换参数对干旱的响应存在显著差异。随干旱程度加剧,第1 年碧玉杨Pn显著(P<0.05)下降,与正常供水相比分别下降了32.15%、44.81%;Gs,Ls,Tr和WUE也均呈下降趋势,而Ci与正常供水无显著差异(P>0.05)。第2年碧玉杨中度、重度干旱与正常供水相比,Pn显著(P<0.05)提高了31.31%、32.37%,Gs与Tr与正常供水无显著变化,重度干旱下Ci显著(P<0.05)下降,而Ls和WUE呈上升趋势并与第1年的变化规律相反(图1)。

图1 干旱对碧玉杨扦插苗功能叶气体交换参数的影响Fig.1 Effects of soil drought on leaf gas exchanges of poplar cuttings
2.2 干旱对碧玉杨生长与生物量分配的影响
干旱显著影响碧玉杨植株生长过程。随干旱程度加剧第1年干旱处理碧玉杨单叶面积逐渐减小而比叶重增大(P<0.05);第2 年中度干旱碧玉杨单叶面积、比叶重与对照相比无显著差异(P>0.05),而重度干旱下分别降低了39.11%、12.39%。干旱显著降低(P<0.05)了碧玉杨的生物量积累,除第2 年中度干旱下茎干重有所提高外,干旱下各器官干重、植株总干重均显著降低(P<0.05);碧玉杨株高、根冠比均显著下降(P<0.05)(表1)。与第1 年相比,第2年正常供水碧玉杨叶干重占比减小、根干重占比增加(P<0.05);干旱下根的生物量分配比例显著降低(P<0.05),而茎的显著增大(P<0.05)(图2)。
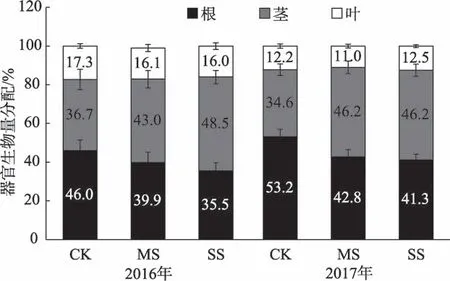
图2 干旱对碧玉杨扦插苗器官间生物量分配的影响Fig.2 Effects of soil drought on organ biomass fractions of poplar cuttings
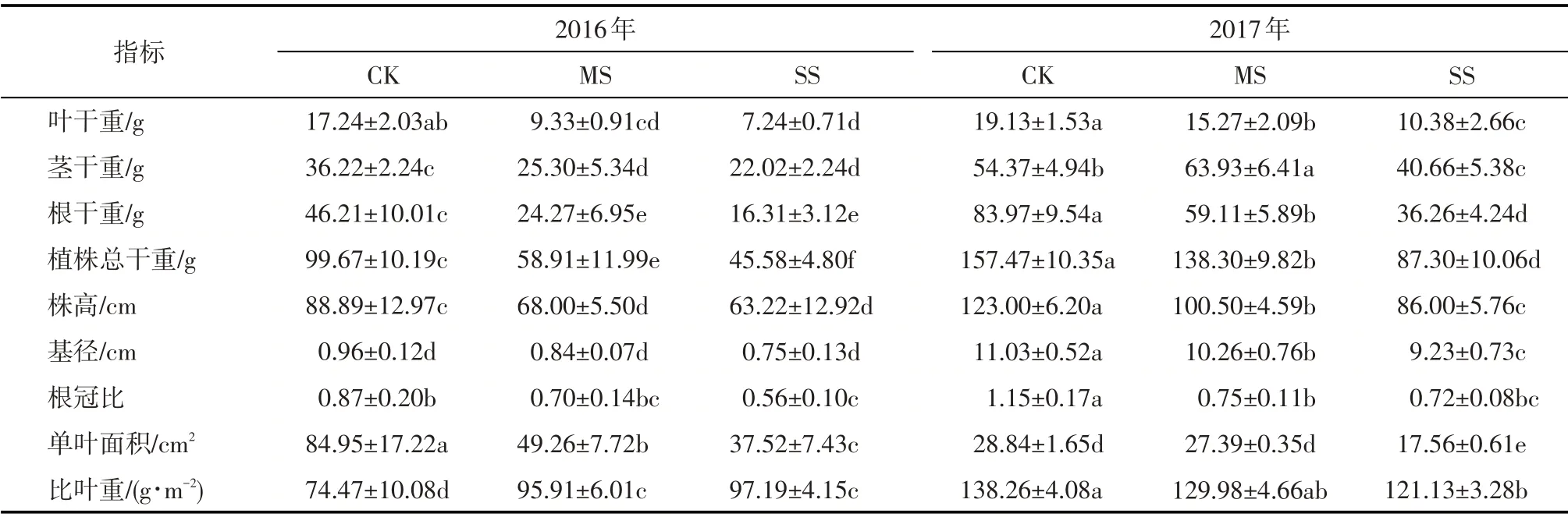
表1 干旱对碧玉杨扦插苗生长指标的影响Tab.1 Effects of soil drought on the growth of poplar cuttings
2.3 干旱对碧玉杨叶片碳氮含量与氮利用的影响
干旱下碧玉杨叶氮含量显著减小(P<0.05);而单位叶面积氮含量有所增加(P<0.05),第1 年重度干旱提高了20.0%(表2)。第1 年干旱下碧玉杨PNUE显著(P<0.05)下降,而碳含量、碳氮比、NUE无显著变化。随干旱时间延长,叶片碳氮含量关系发生了改变,第2 年干旱下碧玉杨叶碳氮含量提高(P<0.05),碳氮比显著降低(P<0.05),单位叶面积氮含量和PNUE无显著变化,中度干旱下NUE显著增加(P<0.05)。2 a各处理的结果回归分析表明,叶片WUE和PNUE之间存在极显著(P<0.01)正相关关系(图3)。

图3 碧玉杨扦插苗叶片瞬时水氮利用的关系Fig.3 Relationship between leaf WUE and PNUE in poplar cuttings
2.4 干旱对根茎叶器官非结构性碳含量的影响
碧玉杨叶、茎和根中的非结构性碳含量依次降低。土壤干旱处理2 a后,叶中的可溶性糖含量显著(P<0.05)降低,重度干旱下茎的可溶性糖含量显著(P<0.05)增加,而根的无显著变化;根茎叶中的淀粉含量各处理间无显著差异;所以,总非结构性碳(TNSC)的变化趋势与可溶性糖含量一致(图4)。图4中结合器官生物量进一步发现茎的可溶性糖总量和TNSC总量分配比例随干旱程度呈逐渐增大趋势,而叶和根的则呈逐渐减小趋势(图5)。
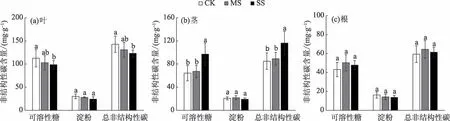
图4 干旱2 a后碧玉杨扦插苗器官非结构性碳含量变化Fig.4 Organ non-structure carbohydrate content changes in poplar cuttings after two years of drought

图5 干旱2 a后碧玉杨扦插苗非结构性碳分配变化Fig.5 Organ non-structure carbohydrate allocation changes in poplar cuttings after two years of drought
3 讨论
3.1 干旱对碧玉杨叶光合作用及水氮利用的影响
干旱对树木叶片光合作用的抑制程度受干旱程度、持续时间以及树种等因素的影响,且可初步分为气孔限制和非气孔限制2种类型[23]。本试验中第1 年干旱下碧玉杨Pn持续降低,而Ls和Ci与正常供水无显著差异(图1),表明其Pn受到非气孔限制,原因可能是叶肉组织光合系统受损、以及CO2从气孔下腔到光合位点的传导度降低[24]。干旱下有些树木的叶片通常会关闭气孔,进而降低Tr并提高WUE[15,24],而本研究中干旱下碧玉杨WUE显著下降,这与中度和重度干旱下107 杨(P.×euramericana‘Neva’)[25]和群众杨[14]的WUE变化趋势一致。同时,碧玉杨叶面积减小、比叶重增加(表1),这有助于维持单位叶面积氮含量不变甚至增加,但却导致PNUE显著降低(表2)。维持较高的叶氮含量对光合作用至关重要,相比于群众杨[14],干旱下碧玉杨具有较强维持叶氮水平的能力,然而,由于干旱下黑杨派杨树的叶肉组织膜系统更容易被破坏[25],导致碧玉杨CO2的叶肉导度显著降低,进而不得不牺牲叶面积以维持较高的Gs和Ci,因此蒸腾耗水较大并导致WUE降低。所以,与树种间的WUE和PNUE通常呈权衡关系[26]不同,干旱下碧玉杨CO2的叶肉导度降低很可能是导致WUE和PNUE均下降的关键原因,这可与Warren 等[24]的研究结果相符合,然而还需要进一步开展抗氧化酶活性和叶肉导度等方面的试验测定进行验证。
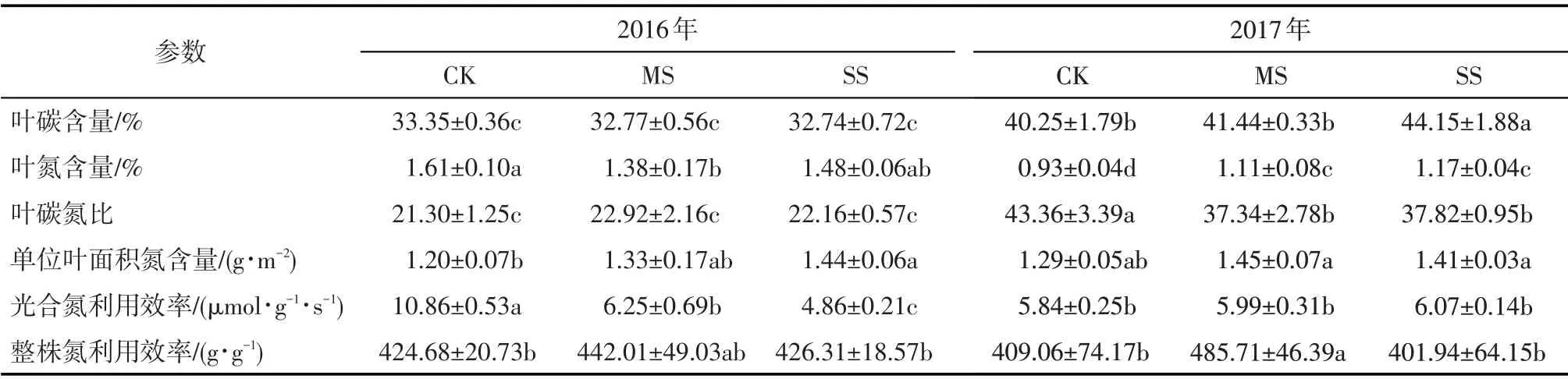
表2 干旱对碧玉杨扦插苗叶片碳氮含量及氮利用的影响Tab.2 Effects of soil drought on the C and N contents and N use of poplar cuttings
与第1 年相比,第2 年正常供水的单叶面积减小比叶重增大,而单位叶面积氮含量维持不变,Gs、Ci和Pn均显著下降;中度、重度干旱下碧玉杨虽叶面积显著减小但Pn小幅提高,中度干旱还显著提高了NUE,这些结果表明二年生碧玉杨幼苗叶片抗旱性比第1 年有所增强,这可能与长期干旱下碧玉杨进一步提高叶片碳投资维持高的叶氮含量有关。
3.2 干旱下碧玉杨器官同化物分配与抗旱性的关系
光合同化物分配过程可影响植物叶片光合速率、茎干运输与根系吸收功能[27]。本试验发现中度和重度干旱下碧玉杨茎的干物质和非结构性碳比例显著增加,而根的显著下降(图2 和图5),这与大多数草本植物和部分树木,如小叶杨[15]和毛白杨(P. tomentosa)[28],干旱下根冠比增大的结果不同。干旱环境中树木茎碳投资增加通常是由于木质部建成的碳消耗增加,比如导管直径减小而导管壁厚度增大[29],可溶性糖等非结构性碳含量增加(图4和图5)。但该现象的形成机制和生理作用还有很大争议,有研究认为是光合产物在叶片中停留时间增长、韧皮部运输速度下降而被动积累[30],大多数研究认为是导管直径减小而壁增厚可降低导管空穴化风险有利于提高木质部的输水安全性[31],可溶性糖积累可通过渗透调节促进导管栓塞修复并维持水分运输[32]。但不论何种解释,结果是中度和重度干旱下碧玉杨根系的生物量和非结构性碳的分配比例均显著降低,这与植物生长的最优分配理论不一致,且明显不利于碧玉杨对水分和氮素等资源的吸收。相比于干旱下根冠比增加的小叶杨[15],虽然碧玉杨叶面积减小以降低蒸腾耗水,但同时也降低了光合固碳的叶面积,导致整株生物量积累大幅降低,且干旱下叶片水氮利用能力均有所下降,根系同化物分配比例更小,因此碧玉杨的整体抗旱性很差,在干旱半干旱区种植碧玉杨后形成“小老树”退化林分的可能性会更大。
4 结论
(1)干旱下一年生碧玉杨幼苗叶片变小变厚,维持较高的叶氮含量,但由于受到非气孔限制,光合能力和瞬时水氮利用能力均显著降低。
(2)二年生碧玉杨叶面积更小而碳氮比和比叶重增加,有助于提高其抗旱性;中度和重度干旱下仍通过减小叶面积维持叶氮含量、光合能力和瞬时水氮利用效率,且中度干旱提高了整株氮利用效率。
(3)土壤干旱下碧玉杨将更多的干物质和非结构性碳分配向茎器官,导致根冠比降低,这虽有助于维持茎的水分运输安全性,但不利于根系的水氮吸收和整株抗旱性。
(4)研究结果初步表明碧玉杨抗旱性较差,不适宜在干旱半干旱地区大面积造林。
猜你喜欢
青年文学家(2021年28期)2021-11-02 18:43:30
宝藏(2019年4期)2019-04-18 08:18:32
文苑(2018年20期)2018-11-09 01:36:00
民族音乐(2018年1期)2018-04-18 03:38:07
浙江农业学报(2017年1期)2017-05-17 06:13:36
西南农业学报(2016年4期)2016-05-17 05:42:13
西南农业学报(2016年4期)2016-05-17 05:42:12
山西农经(2015年7期)2015-07-10 15:00:57
新疆农垦科技(2014年3期)2014-02-28 19:19:28
新疆农垦科技(2014年2期)2014-02-28 19:19:14
