
茶翅蝽成虫对不同植物种类的选择偏好性
2022-06-07 11:31徐元潘洪生修春丽杨益众陆宴辉
植物保护 2022年3期
徐元 潘洪生 修春丽 杨益众 陆宴辉

摘要 茶翅蝽是一种重要的农林害虫,近年来在我国的发生为害程度呈现加重趋势。本文通过田间小区试验和室内行为研究,评价了茶翅蝽成虫对不同植物种类的选择偏好程度。在田间25科115种植物中,84种植物上调查到茶翅蝽成虫,种群发生密度整体较低,但寒麻、芝麻、红麻、籽粒苋、向日葵、大豆、白花菜等植物上成虫密度相对偏高。在室内Y型嗅觉仪测试中,与空白对照相比,茶翅蝽雌雄成虫对苗期和花期的向日葵、大豆、白花菜植株气味均表现出了明显的趋好行为。上述研究结果为深入解析我国茶翅蝽区域分布与发生消长规律提供了科学依据。
关键词 茶翅蝽;成虫密度;寄主偏好;行为选择;植物挥发物
中图分类号: Q968.1
文献标识码: A
DOI: 10.16688/j.zwbh.2021162
Abstract Halyomorpha halys (Stl) is an important pest in agriculture and forestry, and its occurrence and damage have been increasing in recent years. In this paper, we evaluated the host preference of H.halys adults to different plant species by field-plot trial and laboratory behavioral bioassay. Among 115 plant species from 25 families in field plots in 2014 and 2016, H.halys adults were found on 84 host plants, and the population densities of H.halys adults were generally low, but the population abundance of H.halys adults on Cannabis sativa was the highest, followed by Sesamum indicum, Hibiscus cannabinus, Amaranthus hypochondriacus, Helianthus annuus, Glycine max, and Cleome gynandra. Meanwhile, the adults showed obvious preference to three host plants (i.e., H. annuus, G. max and C. gynandra) at both seedling and flowering stages when compared with the blank control in Y-tube bioassay. These results provide scientific basis for further analysis of the regional distribution and occurrence pattern of H.halys in China.
Key words Halyomorpha halys;adult density;host preference;behavioral choice;plant volatile
茶翅蝽Halyomorpha halys (Stl) 属半翅目蝽科,是一种世界性多食性害虫,可为害果树、蔬菜、粮油作物、林木等[1-3]。茶翅蝽成虫和若虫均可取食为害,以其刺吸式口器刺入果实、植物枝条和嫩叶吸取汁液。在果园,果实被害后轻则会呈现部分凹陷斑,重则可造成果实畸形,严重影响水果的品质和质量[4-5]。严重发生时,梨果被害率可达30%~100%,桃果被害率在50%以上,苹果在10%~40%[6]。2010年,茶翅蝽在美国为害导致苹果产业损失高达3 700万美元[7]。
茶翅蝽在中国、朝鲜、韩国和日本属于原发性种类[8],除了在新疆和青海没有报道外,茶翅蝽在我国其他地区均有发生[1,9]。20世纪90年代以来,茶翅蝽相继传入美国、加拿大和欧洲多国,并有继续扩散成为世界性害虫的趋势[2,10]。因此,茶翅蝽在国际上引起了广泛关注,研究人员在该害虫的分类、分布、为害、生物学、生态学习性及生物防治等方面开展了大量工作[2,11-14]。作为一种多食性害虫,茶翅蝽的寄主选择行为、寄主适应能力是研究重点[8,15-17],为茶翅蝽发生防治提供了重要的基础信息。Bergmann等[18]在美國马里兰州通过连续3年试验系统调查了254种木本植物上茶翅蝽发生情况,从中确认了88种寄主植物,并发现被子植物上卵、若虫、成虫密度均显著高于裸子植物上。
本研究通过田间小区试验和室内行为测定,分析了茶翅蝽成虫对我国农田生态系统中常见植物(以草本植物为主)的选择偏好程度,以期为深入解析我国茶翅蝽区域性发生规律提供重要信息。
1 材料与方法
1.1 田间小区试验
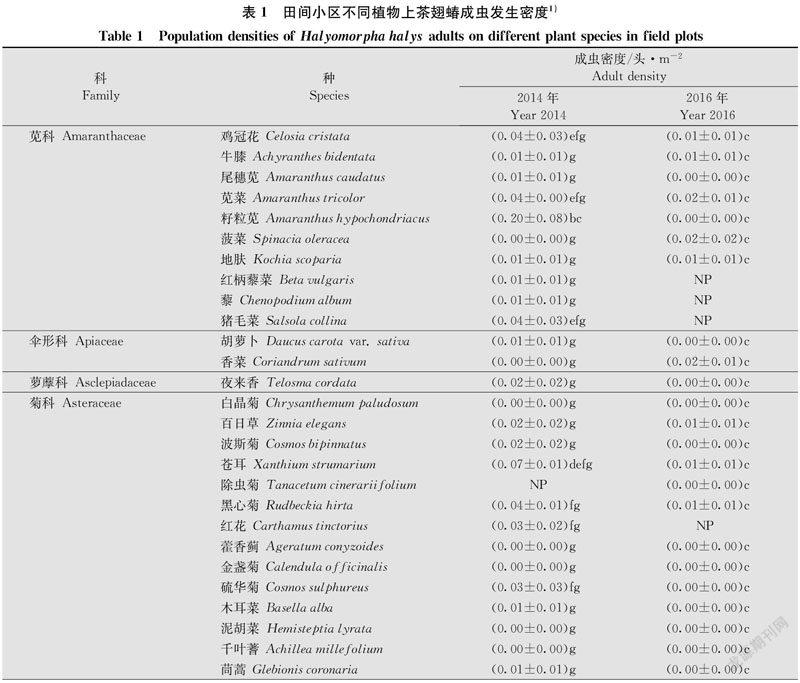
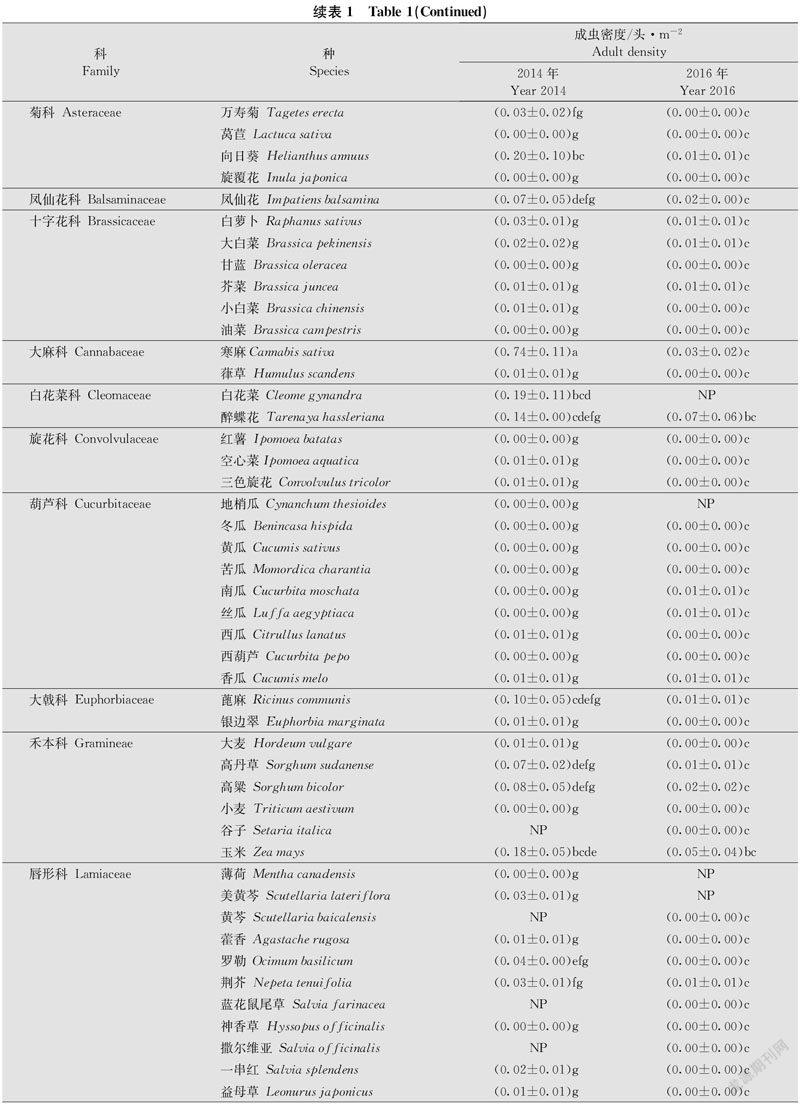
小区试验于2014年、2016年在河北省廊坊市中国农业科学院植物保护研究所廊坊科研中试基地(39.53°N, 116.70°E)开展,共种植25科115种植物(表1),植物种类、小区布置、调查方法同吴月坤等[19]。每种植物每年种植3个小区,小区面积为16 m2(4 m × 4 m),相邻小区间隔 1 m,所有种植小区随机排列。每年5月中旬播种,整个试验过程中不施用任何化学农药。6月下旬至9月中旬,利用目测法对各小区不同植物上茶翅蝽成虫的种群数量进行系统调查。每5 d(2014年)或7 d(2016年)调查一次,每个小区每次随机调查4点,每点调查1 m ×1 m。2014年、2016年分别调查14次、13次。茶翅蝽成虫的种类鉴定参考何振昌等[20]和张金平等[1]。02268ECD-DAD8-4907-A172-11582674779C
1.2 室内行为测定
茶翅蝽成虫从中国农业科学院植物保护研究所廊坊科研中试基地的农田采集。饲养于边长为20 cm的养虫盒中,用玉米和四季豆混合进行继代饲养。养虫室温度为(25±2)℃、相对湿度(60±5)%、光照周期L∥D=14 h∥10 h。饲养1~3代后,2~5日龄茶翅蝽成虫用于选择行为反应试验。
向日葵Helianthus annuus、大豆Glycine max、白花菜Cleome gynandra均为田间小区种植,采用常规的农事管理措施,整个生育期不施用任何化学农药。供试植株用80目的防虫网罩住,以防止茶翅蝽等昆虫为害。
应用Y型嗅觉仪测试茶翅蝽成虫对3种寄主植物的选择行为反应。对于每种植物,苗期处理设置为植物枝条vs.空白,花期处理设置为植物枝条+花vs.空白,选取2~5日龄成虫并饥饿4 h后用于试验,雌雄成虫各测试60头。茶翅蝽成虫寄主选择行为反应测试的Y型嗅觉仪组装及测试方法参照Pan等[21]。试验前,先用清水清洗寄主植物枝条或带花枝条,剪取适宜长度后(鲜重15 g±0.5 g),伤口处用湿润的脱脂棉包裹,然后放入锥形瓶内作为气味源,纯净空气为空白对照。Y型嗅觉仪的气流流速设定为0.4 L/min。将茶翅蝽成虫放入Y形管的主管端,当茶翅蝽成虫越过主管的1/2处开始计时。当茶翅蝽成虫越过两个侧臂的1/3处及以上,记为有反应,若5 min后该成虫仍停留在主管内或未越过侧管的1/3处,则记为无反应。每头成虫只测试1次,每測试2头交换侧管方向及气体方向,每测试4头更换Y形管。每次测试完用无水乙醇清洗Y形管、样品瓶及连接胶管并晾干,以消除装置中残留气味的影响。此试验在温度为(25±2)℃、相对湿度为(60±5)%的实验室内进行,Y型嗅觉仪放置在100 cm × 100 cm × 60 cm的暗室中,嗅觉仪上方40 cm处挂有1盏40 W的日光灯。每次试验开始前,用手持照度计(型号LX1010B,上海均达仪器公司生产)测量嗅觉仪上方的光照强度,确保两臂光照强度相同,避免光照差异对成虫选择行为产生影响。
1.3 数据统计分析
根据2014年、2016年调查次数,分别统计各年度茶翅蝽成虫整个生长季节在不同植物上的发生密度(单位面积上虫量)平均值,采用二因素重复方差分析法(PROC,GLM)比较不同植物种类上的茶翅蝽成虫密度差异。其中,植物种类、年度及其交互作用为固定效应(fixed effects),小区数(即重复数)为随机效应(random effect),多重比较用 SNK(Student-Newman-Keuls)法。所有统计分析均采用SAS 9.3软件进行[22]。
行为反应测试中,首先用选择某一方向的茶翅蝽成虫个体数除以测试中做出选择的昆虫个体总数,即得其对这一方向挥发性气味的选择率。然后利用卡方(χ2)测验法检验茶翅蝽成虫在两个味源间是否呈H0为50∶50的理论分布,计算χ2值和相应的显著性水平P值。未做出选择反应的茶翅蝽成虫不列入统计分析。所有数据均采用SPSS 25.0软件进行分析。
2 结果与分析
2.1 不同植物上茶翅蝽成虫发生密度
在供试的115种植物中,两年中发现有茶翅蝽成虫的植物达84种,其余31种未调查到茶翅蝽成虫。2014年、2016年,田间茶翅蝽发生数量较低,分别调查到成虫289头和107头。广义线性模型(GLM)分析表明,茶翅蝽成虫密度在不同植物种类上(F = 11.17,df = 114 430,P < 0.000 1)、不同年度间(F = 95.14,df = 1 430,P < 0.000 1)均存在显著差异,同时植物种类与年度的交互作用也有显著差异(F = 8.31,df = 99 430,P < 0.000 1)。
2014年,寒麻Cannabis sativa上茶翅蝽成虫平均密度最高,为0.74头/m2,显著高于其他植物种类。其后,依次为红麻Hibiscus cannabinus、籽粒苋Amaranthus hypochondriacus、向日葵、大豆、白花菜、玉米Zea mays、含羞草Mimosa pudica、醉蝶花Tarenaya hassleriana、芝麻Sesamum indicum、蓖麻Ricinus communis,密度为0.26~0.10头/m2。2016年,芝麻上茶翅蝽成虫密度(0.32头/m2)显著高于其他作物,随后依次为大豆、醉蝶花、玉米、寒麻、陆地棉Gossypium hirsutum、红麻,密度范围为0.08~0.03头/m2(表1)。
2.2 茶翅蝽成虫对植物挥发物的行为反应
与空白对照相比,茶翅蝽雌、雄成虫对向日葵、大豆、白花菜苗期植株的枝条均表现出明显的选择偏好性(P < 0.05),且雌、雄成虫对同种植物的选择行为反应无显著差异(P > 0.05)(图1a)。花期的结果与苗期一致,茶翅蝽雌、雄成虫对向日葵、大豆、白花菜的带花枝条均表现出明显的选择偏好性(P < 0.05),且雌、雄成虫对同种植物的选择行为反应间无显著差异(P > 0.05)(图1b)。
3 讨论
茶翅蝽的寄主植物范围广泛,因此在农田生态系统中有着比较复杂的虫源关系。我国之前主要报道茶翅蝽对一些果树等多年生植物的取食为害[3,8],本研究首次记载了多种一年生植物上茶翅蝽的成虫发生,其中很多植物在各地广泛存在,可能在茶翅蝽区域性种群发生中发挥着重要作用。茶翅蝽成虫具有飞行扩散能力,因此发现有成虫存在不足以证明该种植物就是茶翅蝽的寄主植物,但成虫数量多少、密度高低能有效反映茶翅蝽对不同植物的喜好性差异。本研究将有助于对我国茶翅蝽区域性虫源关系与发生规律的深入认识。
诱集植物是一种重要的害虫防治技术,主要是利用害虫成虫对个别寄主植物的偏好性,通过合理种植这些植物吸引害虫来保护主栽作物免受为害[23-25]。例如,绿盲蝽Apolygus lucorum成虫偏好选择绿豆植株, 棉田周边种植绿豆诱集带可明显降低棉花上绿盲蝽种群发生数量[26]。本研究发现茶翅蝽成虫在不同种类植物上的种群数量不同、偏好性各异,其中寒麻、芝麻、红麻、籽粒苋、向日葵、白花菜等植物上茶翅蝽成虫密度较高,是否可作为不同作物田茶翅蝽诱集植物,有待进一步系统评估。目前,向日葵已被用作茶翅蝽诱集植物,在辣椒田周围种植来诱捕茶翅蝽成虫[27]。Nielsen 等[28]研究发现,高粱和向日葵组成的诱集植物对包括茶翅蝽在内的多种蝽具有很好的诱捕作用。02268ECD-DAD8-4907-A172-11582674779C
嗅觉在植食性昆虫寻找、定位和选择寄主植物过程中常发挥着重要作用。李鑫等[29]研究发现,饥饿后的茶翅蝽对食物反应敏感,触角是其嗅食、确定食物方位的主要器官。李秋霖等[30]对茶翅蝽非典型气味受体基因进行了克隆,对其生物信息学特征和组织特异性表达进行了相应分析。本研究在田间调查发现向日葵、大豆、白花菜植株上均有着相对较高的茶翅蝽成虫密度,同时室内行为测定表明嗅觉在茶翅蝽成虫对这三种植物选择行为中具有重要作用,两部分结果高度一致并相互印证。后续有待进一步利用电生理学、行为学等化学生态学研究技术与方法,从茶翅蝽成虫高密度发生的植物种类中分析鉴定引起茶翅蝽成虫触角电生理反应、行为偏好的挥发性物质活性组分,这将对茶翅蝽成蟲的植物源引诱剂以及行为调控措施的创新研发具有重要意义[2]。
参考文献
[1] 张金平, 张峰, 钟永志, 等. 茶翅蝽及其生物防治研究进展[J]. 中国生物防治学报, 2015, 31(2): 166-175.
[2] LESKEY T, NIELSEN A L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management [J]. Annual of Review Entomology, 2018, 63: 599-618.
[3] 徐庆宣, 王甦, 郭晓军, 等. 桃园茶翅蝽的发生危害与防治研究进展[J]. 环境昆虫学报, 2020, 42(4): 877-883.
[4] 余春林, 金新富. 茶翅蝽和斑须蝽对梨树的危害及防治研究[J]. 中国果树, 2002 (2): 5-7.
[5] MAISTRELLO L, VACCARI G, CARUSO S, et al. Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in northern Italy [J]. Journal of Pest Science, 2017, 90(4): 1231-1244.
[6] 李鑫, 龚建宁, 李海林. 茶翅蝽取食行为研究初探[J]. 河北林学院学报, 1996, 11(S1): 230-234.
[7] LESKEY T C, SHORT B D, BUTLER B R, et al. Impact of the invasive brown marmorated stink bug, Halyomorpha halys (Stl), in mid-Atlantic tree fruit orchards in the United States:case studies of commercial management [J/OL]. Psyche: A Journal of Entomology, 2012: 535062. DOI: 10/1155/2013/535062.
[8] LEE D H, SHORT B D, JOSEPH S V, et al. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea [J]. Environmental Entomology, 2013, 42(4): 627-641.
[9] 王浩建, 刘国卿. 秦岭西段及甘南地区昆虫[M]. 北京: 科学出版社, 2005: 279-292.
[10]BARISELLI M, BUGIANI R, MAISTRELLO L. Distribution and damage caused by Halyomorpha halys in Italy [J]. EPPO Bulletin, 2016, 46(2): 332-334.
[11]LEE D H, LESKEY T C. Flight behavior of foraging and overwintering brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae) [J]. Bulletin of Entomological Research, 2015, 105(5): 566-573.
[12]KRITICOS D J, KEAN J M, PHILLIPS C B, et al. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity [J]. Journal of Pest Science, 2017, 90(4): 1033-1043.
[13]KUHAR T P, KAMMINGA K. Review of the chemical control research on Halyomorpha halys in the USA [J]. Journal of Pest Science, 2017, 90(4): 1021-1031.
[14]陈菊红, 米倩倩, 陈立玲, 等. 基于文献计量学的茶翅蝽国内外研究现状分析[J]. 中国植保导刊, 2019, 39(12): 28-36.02268ECD-DAD8-4907-A172-11582674779C
[15]ACEBES-DORIA A L, LESKEY T C, BERGH J C. Development and comparison of trunk traps to monitor movement of Halyomorpha halys nymphs on host trees [J]. Entomologia Experimetalis et Applicata, 2016, 158(1): 44-53.
[16]BOSCO L, NARDELLI M, TAVELLA L. First insights on early host plants and dispersal behavior of Halyomorpha halys (Hemiptera: Pentatomidae) from overwintering to crop colonization [J/OL]. Insects, 2020, 11: 866. DOI: 10.3390/insects11120866.
[17]HOLTHOUSE M C, SPEARS L R, ALSTON D G. Urban host plant utilisation by the invasive Halyomorpha halys (Stl) (Hemiptera, Pentatomidae) in northern Utah [J]. NeoBiota, 2021, 64: 87-101.
[18]BERGMANN E J, VENUGOPAL P D, MARTINSON H M, et al. Host plant use by the invasive Halyomorpha halys (Stl) on woody ornamental trees and shrubs [J/OL]. PLoS ONE, 2016, 11(2): e0149975. DOI: 10.1371/journal.pone.0149975.
[19]吴月坤, 刘冰, 潘洪生, 等. 小花蝽在不同植物上的种群密度[J]. 中国生物防治学报, 2019, 35(4): 527-535.
[20]何振昌, 张治良, 黄峰, 等. 中国北方农业害虫原色图鉴[M]. 沈阳:辽宁科学技术出版社, 1997.
[21]PAN Hongsheng, LU Yanhui, XIU Chunli, et al. Volatile fragrances associated with flowers mediate host plant alternation of a polyphagous mirid bug [J/OL]. Scientific Reports, 2015, 5: 14805. DOI: 10.1038/srep14805.
[22]SAS Institute. SAS/STAT9.3 users guide [M]. Cary, North Carolina: SAS Institute Inc, 2011.
[23]FOSTER S P, HARRIS M O. Behavioral manipulation methods for insect pest-management [J]. Annual Review of Entomology, 1997, 42: 123-146.
[24]SHELTON A M, BADENES-PEREZ F R. Concepts and applications of trap cropping in pest management [J]. Annual Review of Entomology, 2006, 51: 285-308.
[25]陆宴辉, 张永军, 吴孔明. 植食性昆虫的寄主选择机理及行为调控策略[J]. 生态学报, 2008, 28(10): 5113-5120.
[26]LU Yanhui, WU Kongming, WYCKHUYS K A G, et al. Potential of mungbean, Vigna radiatus as a trap crop for managing Apolygus lucorum (Hemiptera: Miridae) on Bt cotton [J]. Crop Protection, 2009, 28(1): 77-81.
[27]SOERGEL D C, OSTIGUY N, FLEISCHER S J, et al. Sunflower as a potential trap crop of Halyomorpha halys (Hemiptera, Pentatomidae) in pepper fields [J]. Environmental Entomology, 2015, 44(6): 1581-1589.
[28]NIELSEN A L, DIVELY G, POTE J M, et al. Identifying a potential trap crop for a novel insect pest, Halyomorpha halys (Hemiptera: Pentatomidae), in organic farms [J]. Environmental Entomology, 2016, 45(2): 472-478.
[29]李鑫, 尹翔宇, 馬丽, 等. 茶翅蝽的行为与控制利用[J]. 西北农林科技大学学报(自然科学版), 2007, 35(10): 139-145.
[30]李秋霖, 赵吉兰, 田嘉慧, 等. 茶翅蝽非典型气味受体基因的克隆及其生物信息学特征和组织特异性表达分析[J]. 植物保护学报, 2020, 47(6): 1219-1227.
(责任编辑:田 喆)02268ECD-DAD8-4907-A172-11582674779C
