长链非编码RNA LUCAT1在恶性肿瘤中作用及机制的研究进展
2022-06-03 02:35:54白易综述李仲珉张雅敏审阅南开大学医学院附属医院天津市第一中心医院肝胆外科天津3009天津医科大学一中心临床学院天津300070
中国肿瘤生物治疗杂志 2022年3期
白易 综述;李仲珉,张雅敏 审阅(.南开大学医学院附属医院 天津市第一中心医院 肝胆外科,天津 3009;.天津医科大学 一中心临床学院,天津 300070)
恶性肿瘤是一种威胁人类健康的严重疾病。多年来,对肿瘤生物学的研究集中在蛋白质编码基因的参与上,而长链非编码RNA(long non-coding RNA,lncRNA)是一种长度超过200个核苷酸且不具备蛋白质编码功能的RNA 分子,曾被认为是转录垃圾,近30 年才发现其在细胞活动中起到关键的调控作用[1-2]。目前认为,lncRNA 具有高度的组织和细胞特异性,其来源复杂、结构多样,可以在多个维度调控DNA、RNA 和蛋白质的功能[3-6]。位于5 号染色体q14.3 区的反义链的肺癌相关转录物1(lung cancerrelated transcript 1,LUCAT1)最早是在吸烟的肺癌患者中被发现的[7-8]。越来越多的研究结果[9]发现,LUCAT1 在多种类型恶性肿瘤中具有促进肿瘤发生和发展的作用,并与患者的肿瘤大小、组织学分级、TNM分期和生存期(OS)等多项临床特征显著相关。本文综述了LUCAT1 促进肿瘤发生和发展的作用机制、在不同类型恶性肿瘤(主要为消化系统和泌尿生殖系统)中的表达和功能、在预后评估中的研究进展,旨在为恶性肿瘤的早期诊断和治疗及预后评估提供参考资料。
1 LUCAT1促进肿瘤发生和发展的作用及机制
1.1 与DNA的相互作用
抑癌基因的启动子区甲基化会导致转录沉默和基因功能丧失,进而促进肿瘤的形成和发展。研究结果[10]表明,LUCAT1可以通过甲基化、磷酸化、乙酰化或泛素化等转录后机制增强DNA甲基转移酶1的表达,维持抑癌基因启动子区甲基化,并下调其表达,促进肿瘤的形成。多梳抑制复合物2 由EZH2、SUZ12 和EED 组成,能催化组蛋白3 上的第27 位赖氨酸的三甲基化(H3K27me3),继而调控下游基因表达。有研究结果[11-12]显示,LUCAT1 可以招募并结合EZH2,随后与抑癌基因p21 和p57 的启动子区相结合,下调其表达并促进肿瘤细胞增殖。
1.2 与miRNA的相互作用
miRNA可以与靶基因mRNA的3'UTR相互作用,使mRNA 去甲基化和失稳。作为竞争性内源RNA(competitive endogenous RNA,ceRNA),lncRNA可以像分子海绵一样在细胞质中吸附miRNA并降低其活性,间接上调其下游靶基因的表达。多项研究结果[13]表明,LUCAT1可以作为ceRNA调控不同的miRNA及其靶基因(表1)。在肝母细胞瘤中,LUCAT1通过与miR-301b竞争性结合,上调STAT3的表达,促进肝母细胞瘤细胞的增殖、迁移和侵袭。在乳腺癌中,LUCAT1通过吸附miR-5582-3p、miR-7-5p 和miR-181a-5p,上调SOX2、KLF6和KL15的表达,激活Wnt/β-catenin通路,提高乳腺癌干细胞的细胞干性,促进肿瘤生长[14-15]。此外,LUCAT1不仅可以与miR-375结合,间接上调YAP1来促进肾透明细胞癌(clear cell renal cell carcinoma,ccRCC)细胞增殖、迁移和侵袭,在口腔鳞状细胞癌及胶质瘤中也同样具有致癌作用[16-18]。但也有研究结果[19]显示,LUCAT1在肝恶性肿瘤中可以通过吸附miR-181d-5p并降低其表达水平,降低肝恶性肿瘤的侵袭潜力,抑制其复发。
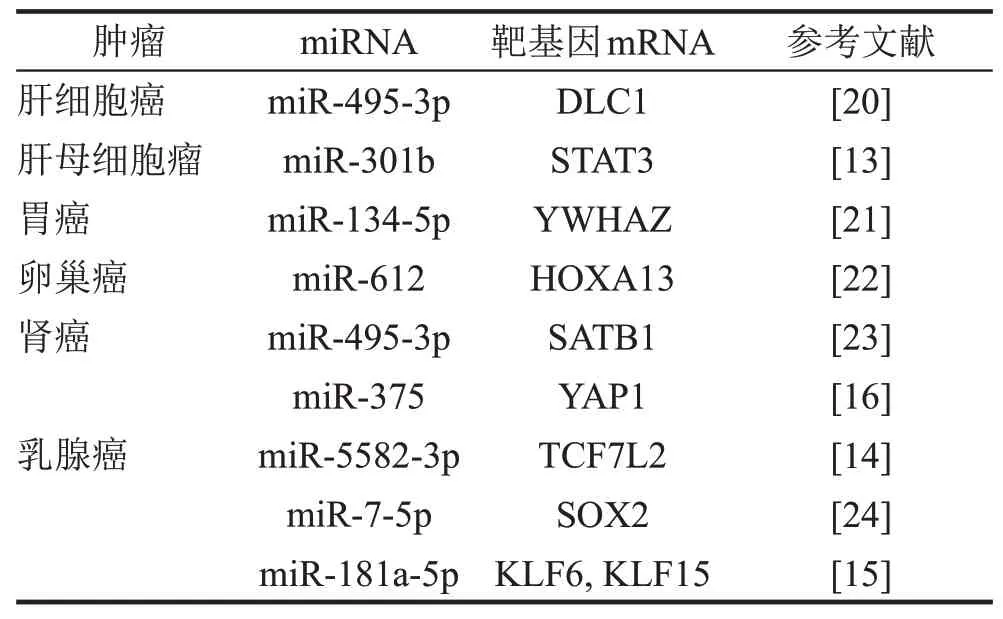
表1 LUCAT1与miRNA结合的不同类型肿瘤靶基因
1.3 与蛋白质的相互作用
由于lncRNA 较长,易于形成空间二级结构,因此也可以通过结合蛋白质参与分子调控,LUCAT1可以和多种蛋白质相互结合发挥作用。如:下调LUCAT1 后,CDK2、CDK4、cyclin D1、cyclin E1 和cyclin E2的表达显著降低,继而降低视网膜母细胞瘤蛋白(retinoblastoma protein,Rb)的磷酸化,增强p21蛋白表达,并诱导细胞周期停滞在G1 期[25-27]。LUCAT1也可以干扰核糖体的生物合成,下调rRNA、18S RNA 和pre-rRNA 的表达,促进结直肠癌(colorectal cancer,CRC)进展[28]。此外,LUCAT1也可以与核仁素直接结合,继而解除核仁素对MYC的抑制作用,促进CRC细胞增殖[29]。LUCAT1还可以与多聚嘧啶区结合蛋白1 相互作用,调控下游靶基因CD44、APP、CLSTN1、MBNL1 和ZNF207 的选择性剪接,阻碍这些靶基因参与的细胞增殖和DNA 损伤修复进程[30]。最后,LUCAT1 还可以抑制膜联蛋白A2 的磷酸化并影响其翻译后修饰,抑制膜联蛋白A2-S100A10 四聚体的降解,加速纤溶酶原向纤溶酶的转化,从而激活金属蛋白酶,促进肝细胞癌(hepatocellular carcinoma,HCC)的进展和转移[31]。
2 LUCAT1在消化系统恶性肿瘤中的作用
2.1 食管癌
有研究结果[10]发现,LUCAT1在食管鳞状细胞癌(esophageal squamous cell carcinoma,ESCC)组织和细胞中表达均显著上调,LUCAT1 高表达的患者OS更短。下调LUCAT1 后,可以抑制癌细胞增殖、侵袭和迁移,并诱导细胞凋亡。进一步机制发现,LUCAT1 主要通过调控DNA 甲基转移酶1 下调抑癌基因的表达,促进ESCC 的进展。因此,LUCAT1 不仅可以作为ESCC的预后生物标志物,还可以作为潜在的治疗靶标。
2.2 胃癌
CHI 等[21]发现,LUCAT1 在胃癌组织中的表达水平显著高于癌旁组织,且高表达与患者的OS和无病生存期(DFS)缩短相关,敲降LUCAT1后可以显著抑制胃癌细胞增殖、迁移和侵袭。在胃癌组织和细胞中,LUCAT1和miR-134-5p的表达呈负相关。既往的研究结果[32-34]表明,miR-134-5p 可以通过抑制肿瘤细胞增殖、迁移和侵袭,在ESCC、HCC、肾癌和乳腺癌等多种肿瘤中发挥抑癌作用。进一步通过TargetScan 在线工具预测发现,miR-134-5p 与YWHAZ基因之间存在结合位点,且YWHAZ在胃癌中的表达与miR-134-5p呈负相关。经实验验证后发现,LUCAT1 可以通过ceRNA 机制竞争性结合miR-134-5p,继而上调YWHAZ mRNA 的表达,促进胃癌细胞的进展[21]。该研究结果提示,LUCAT1不仅具有诊断胃癌、评估其预后的潜质,也是胃癌潜在的治疗靶点。
2.3 HCC
LUCAT1在HCC组织中表达上调已经得到了多项研究[19,31,35]的证实。在HCC异种移植模型中,过表达LUCAT1 显著增强HCC 细胞的增殖、迁移和侵袭能力,并促进体内肿瘤的生长和转移[31]。JIAO等[19]通过回顾性分析371例HCC患者的测序数据和临床数据发现,LUCAT1 高表达与肿瘤分化差、病理分期晚相关,且高表达LUCAT1 患者的OS 更短,多因素分析也证实,LUCAT1 是HCC 预后不良的独立风险因素。因此,LUCAT1 可以作为HCC 治疗的靶点和预后相关的生物标志物。
2.4 胰腺导管腺癌(pancreatic ductal adenocarcinoma,PDAC)
PDAC 是最常见的胰腺癌病理亚型。研究结果[25,36]表明,LUCAT1 在PDAC 组织和细胞中表达均显著上调,LUCAT1高表达与PDAC肿瘤大小和淋巴结转移相关。体外实验结果[36]表明,下调LUCAT1表达可以抑制PDAC 细胞的增殖、迁移和侵袭,并诱导细胞周期停滞。基于肿瘤异种移植模型的体内实验[36]进一步证实,LUCAT1的下调可以显著减小肿瘤质量和体积。通过生物信息学分析和荧光素酶报告基因实验发现,LUCAT1可以作为miR-539的分子海绵发挥促癌作用。此外,LUCAT1 还可以通过诱导AKT和MAPK的磷酸化,促进PDAC细胞发育[25]。
2.5 CRC
据报道[29-30,37-38],CRC 组织和细胞中LUCAT1 的表达同样显著上调,且LUCAT1高表达降低了患者的OS。相关研究结果[28]表明,下调LUCAT1 表达不仅可以显著抑制CRC细胞的增殖、迁移和侵袭能力,诱导细胞凋亡,并增强CRC 细胞对化疗药物奥沙利铂和5-FU 的敏感性。机制研究结果[28]表明,LUCAT1主要通过与UBA52 结合而活化RPL40-MDM2-P53信号通路,促进CRC的发生和发展。
3 LUCAT1在泌尿生殖系统恶性肿瘤中的作用
3.1 肾癌
肾细胞癌(renal cell carcinoma,RCC)是泌尿系统最常见的恶性肿瘤之一,其中ccRCC 是最常见的RCC 亚型,约占所有RCC 的75%。多项研究[9,16,39]报道了LUCAT1同ccRCC患者的不良预后、分型、TNM分期和转移的相关性,但其主要作用机制尚存在争议。XIAO 等[11]研究发现,LUCAT1 的下调可显著抑制肾癌细胞的增殖、迁移和侵袭,并可使癌细胞停滞在G1 期;LUCAT1 在ccRCC 细胞核的表达明显高于细胞质。LUCAT1主要与PRC相互作用,抑制p57的表达,在ccRCC 中发挥癌基因的作用。而WANG等[23]的研究则发现,LUCAT1可作为ceRNA竞争性结合miR-495-3p,从而上调特异性核基质结合区结合蛋白1(special AT-rich sequence binding protein 1,SATB1)的表达[40],SATB1 在多种肿瘤的侵袭和转移中发挥重要作用。ZHENG 等[26]的研究结果则表明,敲降LUCAT1 可以抑制cyclin D1、CDK4 和p-Rb 的表达,诱导细胞周期阻滞于G1期;过表达LUCAT1也可以促进丝氨酸/苏氨酸激酶的磷酸化,抑制糖原合成酶激酶-3β(glycogen synthase kinase-3β,GSK-3β)的活性。趋化因子CXCL2 可以上调LUCAT1 在RCC 细胞中的表达,提示CXCL2/LUCAT1/AKT/GSK-3β轴在RCC的发生和发展过程中起关键作用。因此,LUCAT1 在RCC 中的亚细胞定位及具体分子调控机制仍有待进一步研究。
3.2 卵巢癌
多项研究结果[41-44]证明,HOXA13 同恶性肿瘤的进展密切相关。LUCAT1在卵巢癌组织中显著上调,并与晚期卵巢癌的分期、转移和不良预后呈正相关[22]。同时,发现LUCAT1 可以通过竞争性结合miR-612而上调HOXA13的表达,促进卵巢癌细胞增殖、迁移和侵袭。也有研究结果[45]发现,LUCAT1 的3'UTR含有高度保守的miR199a-5p结合位点。qPCR检测结果进一步证实,LUCAT1的上调可以显著降低miR-199a-5p 的表达,而miR-199a-5p 已被证明可以调控细胞的多种生物学过程,包括增殖、凋亡、迁移和侵袭[46-47]。总之,LUCAT1 是一个非常有前景的卵巢癌的治疗靶点。
3.3 宫颈癌
宫颈癌也是妇科最常见的恶性肿瘤之一,多项研究结果[48]表明,多种lncRNA 在宫颈癌组织和细胞中表达异常,参与多条信号通路的调控,影响宫颈癌细胞的增殖、凋亡、迁移和侵袭等过程。ZHANG等[49]研究发现,核转录因子SP1能上调LUCAT1在宫颈癌细胞中的表达。通过生物信息学工具预测发现,miR-181a 是LUCAT1 的潜在结合miRNA,而荧光素酶报告基因实验和RNA免疫沉淀实验也证实了这一点。值得注意的是,miR-181a 在包括宫颈癌在内的多种类型肿瘤中低表达,具有抑癌作用[50-53]。通过机制研究[49]发现,SP1 可以直接与LUCAT1 的启动子区域结合并激活其转录,然后通过竞争性结合miR-181a促进宫颈癌细胞的增殖、迁移和侵袭。
4 LUCAT1在其他恶性肿瘤中的作用
4.1 胶质瘤
胶质瘤是神经系统中最常见的恶性肿瘤。GAO等[18]研究结果发现,LUCAT1在胶质瘤组织和细胞中表达显著上调,且在肿瘤分期较晚的胶质瘤患者中表达更高,下调LUCAT1表达可显著抑制胶质瘤细胞的增殖和侵袭。利用生物信息学工具DIANA 对LUCAT1 的靶点进行预测发现,miR-375 与其存在潜在的结合位点。通过荧光素酶报告基因实验进一步证实了二者之间存在的结合位点。qPCR 实验结果也证实,miR-375 与LUCAT1 表达呈负相关关系,在胶质瘤组织中表达下调。功能实验证实,miR-375可以逆转LUCAT1 对胶质瘤细胞增殖和侵袭的促进作用。
4.2 肺癌
LUCAT1 在非小细胞肺癌(non-small cell lung cancer,NSCLC)组织中的表达水平显著高于癌旁组织,在肿瘤较大、TNM 分期较晚和预后差的NSCLC患者中其表达水平更高[12]。实验结果[12]证明,敲除LUCAT1基因后显著抑制NSCLC细胞在体外和体内的增殖能力;机制研究表明,LUCAT1 与多梳抑制复合物2 之间存在相互作用,通过形成复合物发挥对p21和p57的表观抑制作用。
4.3 乳腺癌
乳腺癌是全球范围内女性最常见的恶性肿瘤和肿瘤死亡的首要原因[54]。多项研究结果[14-15,24,55-56]发现,LUCAT1 在乳腺癌组织和细胞中表达均显著上调,且与肿瘤大小、淋巴结转移和临床分期密切相关。ZHENG 等[14]研究发现,LUCAT1 可作为miR-5582-3p的分子海绵影响其与转录因子7类似物2 的靶向结合,继而活化Wnt/β-catenin 通路,在体内和体外促进乳腺癌干细胞的自我更新和增殖。此外,LI 等[24]的研究结果发现,LUCAT1 也可以作为miR-7-5p 的分子海绵,间接上调转录因子SOX2,从而促进乳腺癌细胞的增殖、迁移和侵袭。
5 结语
lncRNA是肿瘤发生和发展的重要调控因子。随着研究的不断深入,lncRNA 在肿瘤发生和发展中的作用愈发值得深入探讨和总结。结合现有研究,lncRNA LUCAT1 在多种类型肿瘤中表达上调,并与肿瘤大小、组织学分级、TNM 分期和OS 等临床特征显著相关,是潜在的肿瘤诊断和预后评估生物标志物。值得一提的是,在体内和体外敲降LUCAT1后可显著抑制各种类型肿瘤细胞的增殖、迁移和侵袭能力,表明LUCAT1 是肿瘤免疫治疗的潜在靶点。LUCAT1发挥的分子调控机制与其亚细胞定位有关。在细胞核中,LUCAT1可调控DNA甲基化,并与核蛋白相互作用进行基因调控;在细胞质中,LUCAT1 则可作为基因调控的ceRNA,调控mRNA代谢。总之,本综述总结了LUCAT1 在各种类型肿瘤中的作用及其分子机制,拓宽了对肿瘤表观遗传学发病机制的理解,并为研发新的肿瘤诊断生物标志物和治疗靶点提供了新的思路。
猜你喜欢
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
作文教学研究(2016年1期)2016-07-05 12:22:47
中国卫生标准管理(2015年3期)2016-01-14 03:41:46
磁共振成像(2015年8期)2015-12-23 08:53:14
吉林大学学报(医学版)(2015年5期)2015-12-16 15:43:57
医学研究杂志(2015年9期)2015-07-01 17:28:27
医学研究杂志(2015年8期)2015-06-22 14:00:57
中国当代医药(2015年20期)2015-03-01 02:04:29
中国当代医药(2015年9期)2015-03-01 02:02:00