
RNA解旋酶参与植物叶片衰老调控研究进展
2022-06-01 07:55张战辉张慧李伟亚付志远郭战勇
河南农业大学学报 2022年2期
张战辉, 张慧, 李伟亚, 付志远, 郭战勇
(河南农业大学农学院,河南 郑州 450002)
植物叶片是光合作用的主要器官。叶片衰老是植物长期进化过程中与环境相适应形成的一种生存机制,决定着植物的适应性和产量,受控于复杂、精确的调控网络,外界环境的变化和生物胁迫的发生往往会导致叶片衰老的起始[1]。作物的光合产物是人们的主要食物来源[2],作物生长发育过程中,叶片光合产物和吸收的营养物质不断积累,而衰老过程则是成熟叶片中积累的光合产物和营养物质被转移到正在发育的新叶片和籽粒中的过程,叶片功能期延长能提高作物产量[3]。因此,叶片衰老对于粮食生产具有重要的决定性作用,如C4植物玉米,叶片具有较C3植物更高的光合效率,且生育期较短,叶片早衰会严重影响籽粒产量和品质。同样,玉米叶片衰老也是植株衰老开始的标志,伴随着功能叶全部衰老,茎秆、雌穗和籽粒开始脱水,玉米成熟期的籽粒、雌穗含水量和抗倒性是重要的适宜机收性状,晚熟不利于籽粒的机械化收获。研究叶片衰老分子调控机制不仅具有极其重要的生物学意义,而且具有重要的应用价值。
解旋酶包括DNA解旋酶和RNA解旋酶,是生物中广泛存在的依赖于ATP的核酸解旋酶类,在各种生物过程中发挥着至关重要的作用[4]。DNA解旋酶主要参与DNA复制、重组、修复和转录过程的调控,以及维持染色体的稳定[5]。RNA解旋酶可以解开RNA二级结构中存在的双链,主要作为RNA伴侣协助结合的RNA从细胞核转运出细胞核,进而参与转录、核糖体组装、RNA剪接和mRNA翻译[6]。越来越多的研究表明,RNA解旋酶参与了各植物生长发育过程和非生物胁迫响应的调控[7-9]。本研究首先对模式植物拟南芥和玉米叶片衰老调控研究进行概括,然后对RNA解旋酶的分类和细胞学功能进行阐述,进而对RNA解旋酶参与叶片衰老调控的研究进展进行重点总结,并对玉米遗传改良中的潜在应用进行探讨。旨在为理解植物RNA解旋酶参与的叶片衰老调控机制提供必要的依据和参考,为成熟期玉米叶片的遗传改良提供重要理论依据,并为未来研究指明方向。
1 解旋酶分类与生物学功能
1.1 解旋酶分类
根据氨基酸保守序列,可将解旋酶划分为6个家族(superfamily,SF),即SF1~SF6。SF1和SF2家族包括了所有真核生物RNA解旋酶,SF3~SF5家族主要为病毒和细菌DNA解旋酶,SF6家族主要为MCM(微染色体维持蛋白)解旋酶(图1)。其中,SF2家族成员最为庞大,同时包含真核生物所有的DNA和RNA解旋酶,具有丰富的功能多样性[4]。
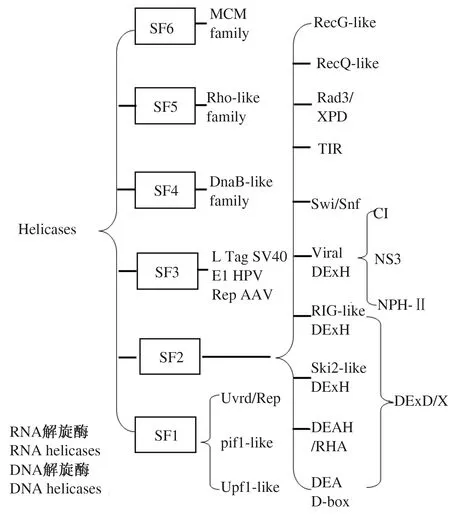
本图根据SERAJ等[4]的研究进行绘制。
植物中DNA解旋酶主要存在于细胞核、线粒体和叶绿体中。相关研究表明,DNA解旋酶参与了许多生物学过程,包括DNA重组、DNA复制、翻译起始、rDNA的转录与前期pre-rRNA加工、双链断裂修复、端粒长度维持、核苷酸切除修复、细胞分裂、DNA甲基化、细胞周期和细胞基础活动[4-5,10-12]。植物RNA解旋酶分布于细胞核、细胞质、叶绿体、线粒体等细胞器中,主要包括DEAD-box、DEAH-box、DExD/H-box和Upf1-like家族,参与mRNA前体剪接加工、rRNA剪接加工、核糖体生物合成、RNA运输、mRNA代谢、mRNA伴侣、mRNA翻译起始、mRNA的表观修饰、miRNA生成、siRNA生成等生物过程[4,6-7,13-15]。前人利用生物信息学方法在拟南芥、水稻、大豆、玉米中分别鉴定到161、149、213和136个RNA解旋酶编码基因,其中多数RNA解旋酶的生物学功能尚需进一步研究[7]。其中,138个玉米RNA解旋酶基因编码215种RNA解旋酶,包括57个DEAD-box,31个DEAH-box,50个DExD/H-boxRNA解旋酶基因,大多数RNA解旋酶在不同组织器官和发育时期呈现差异表达,且部分RNA解旋酶基因转录存在可变剪接[7]。
1.2 RNA解旋酶的生物学功能
细胞内大多数 RNA 分子是单链线状,但具有生物活性的RNA会在某些区域碱基互补配对,而自身折叠形成高度结构化的二级、三级结构。在体内RNA的结构及RNA-蛋白质复合物,即核糖核蛋白(RNP)和一切生物大分子的结构一样是处于动态平衡的过程中,在其功能周期要经历广泛的构象变化。RNA解旋酶可以在ATP的驱动下解开RNA双链,维持单链结构,参与RNA构象变化的调控,从而实现对RNA生物学功能的调控。近年来,植物RNA解旋酶的功能逐渐受到关注,尤其是在作物农艺性状和逆境响应中的调控作用。其中,DEAD-box RNA解旋酶ZmPMH2-5140、AtSMN2、OsRH42、AtUAP56、AtPMH2、AtRID1、AtRCF1,以及Ski2-likeRNA解旋酶ISE2被证明参与mRNA前体剪接过程[16-26];DEAD-box RNA解旋酶SlDEAD39、AtRH50、AtRH36、AtRH18、AtRH57、AtDDX47、AtRH7/PRH75、AtMTR4、OsTOGR1、OsRH17、AtRH22、AtRH3和ZmDRH1参与了rRNA剪接加工与核糖体的生物合成[27-37];DEAD-box RNA解旋酶AtUAP56和THO/TREX组分TEX1被证明参与了mRNA的转运[38-41];Upf1-like家族RNA解旋酶AtUPF1、AtUPF2和AtUPF3,DEA(D/H)-Box家族RNA解旋酶AtDDX3、AtDDX5和AtDDX6被证明参与无义介导的RNA降解途径[8,42-43];DEAD-box RNA解旋酶OsTCD33、OsRH58、SlDEAD31、OsSUV3和AtPRH75被证明参与了RNA代谢的调控[44-48];DEAD-box RNA解旋酶eIF4A和BrRH22被证明参与了mRNA翻译的起始[28,49];此外,eIF4A1(DEAD-box家族)、UPF1(Upf1-like家族)、AtSTRS1(DEAD-box家族)、AtSTRS2(DEAD-box家族),AtRH27(DEAD-box家族)、AtSMA1(DEAD-box家族)分别被证明参与了表观遗传修饰和miRNA的生成途径[50-54]。RNA解旋酶通过以上所述途径参与植物逆境响应[4,50]、受精卵分裂[51]、生长发育[42,49,55]、病原免疫[8]、雌雄配子发育[56]、叶片衰老[9,57]等生物学过程的调控。
1.3 RNA解旋酶参与的植物叶片衰老进程调控
越来越多的研究表明,RNA解旋酶在植物生长发育和非生物胁迫响应中发挥着重要作用,但其调控植物叶片衰老分子机制仅有少量的研究。拟南芥中,一种RNA解旋酶是GVS1调控叶片衰老途径中的重要组分,但该RNA解旋酶的生物学功能尚需进一步研究[57]。AtUPF1的T-DNA插入突变体分别表现出开花期提前、种子增长、持续光照致死、叶片缩小和叶片衰老的表型[9]。水稻中,Upf1-likeRNA解旋酶OsPLS2可能通过无义介导的mRNA降解途径调控叶片的衰老,该RNA解旋酶突变体表现出远红光、H2O2或ABA敏感。水稻DEAD-boxRNA解旋酶PDH45或SUV3的过表达可以降低细胞的凋亡,进而延缓叶片衰老,其参与的叶片衰老调控可能是通过调控端粒酶的活性实现的[58]。目前,尚无RNA解旋酶参与玉米叶片衰老调控的报道,笔者所在课题组通过对玉米叶片早衰突变体进行基因图位克隆,确定突变体基因编码一种人类Smubp-2同源的RNA解旋酶,该RNA解旋酶通过未知机制参与细胞自噬过程的调控,从而控制玉米叶片衰老的起始。
植物叶片衰老过程伴随着叶绿体的降解,叶绿体发育的调控可能与叶片衰老相关。拟南芥中,RNA解旋酶AtISE2、AtRH22、AtRH3分别被证明参与了叶绿体ABA信号转导、RNA剪辑和核糖体亚基的生物合成[21,34-35]。水稻OsTCD33被证明参与了低温调减下叶绿体发育的调控[44],ZmRH3被证明参与了玉米叶绿体核糖体的生物合成[35]。这些RNA解旋酶可能通过调控叶绿体发育和代谢活动控制叶片衰老进程。不同极端外界条件可以诱发叶片的衰老。大量的研究表明,RNA解旋酶在干旱、高盐、高温、低温胁迫响应中发挥着极其重要的作用[18,27,30,44-45,59-61]。这些研究为RNA解旋酶参与植物叶片衰老调控提供了间接证据,尤其是作物中的研究可为相关性状的遗传改良和新品种选育提供必要的理论依据。其中,RNA解旋酶可以通过响应环境因子调控植物叶片衰老的起始,可应用与作物高产和宜机收性状的遗传改良。
2 拟南芥和玉米叶片衰老进程及调控机制研究进展
2.1 拟南芥叶片衰老进程及调控机制
衰老是植物生长发育周期的最后一个阶段,在细胞、组织、器官和植株水平上均表现出明显的衰退迹象,是植物在长期进化中形成的适应环境和生存的一种策略[62]。叶片衰老进程受内因和外因的共同作用,在植物生长发育过程中受不同分子途径的严格调控[63]。导致叶片衰老的外在因素包括干旱、高盐、极端温度、营养、遮阴和病原侵染等,而内源信号主要为植物激素,包括生长素(auxin)、细胞分裂素(cytokinin)、脱落酸(abscisic acid,ABA)、茉莉酸(jasmonic acid,JA)、赤霉素(gibberellic acid,GA)等。外部环境因素和内源信号形成复杂的衰老调控网络(图1)[3,64-65]。除了生物和非生物胁迫,调控叶片衰老的外部因素还包括光照和昼夜节律[66-67]。植物在养分充裕、适宜的环境条件下,叶片衰老主要决定于叶龄和植株的发育进程[68]。细胞学水平,细胞自噬在维持细胞蛋白质的动态平衡中发挥至关重要的作用,细胞自噬的过度和缺少均可导致衰老的起始,是调控叶片衰老的主要途径[69]。
拟南芥中,叶片衰老诱导大量基因的表达发生显著改变,根据其表达类型可以划分为衰老下调基因(senescence down regulated genes,SDGs)和衰老相关基因(表达量上调,senescence-associated genes,SAGs),其中SAGs在植物叶片衰老起始、推进和非生物胁迫响应中发挥着重要的作用[70]。前人利用遗传学、基因组学、转录组学、蛋白质组学和代谢组学分析等方法对植物叶片衰老相关机制进行了解析,鉴定出大量的衰老相关基因[62,71]。叶片衰老过程中,不同内源信号植物激素之间存在互作效应,如生长素和细胞分裂素可以抑制叶片衰老,茉莉酸、脱落酸、水杨酸、赤霉素则促进叶片衰老(图2)[3,72]。而且,叶片衰老受染色质、转录、转录后、翻译和翻译后水平的基因表达调控,不同水平分子调控构成复杂的调控网络[3,73-74]。虽然,模式植物拟南芥叶片衰老已有大量的研究,相关的分子调控机制已得到较为深入的解析,但叶片衰老起始和衰老进程推进的精确控制尚需进一步研究。

根据LI等[71]和WOO等[72]进行本图片绘制。
2.2 玉米叶片衰老研究进展
叶片衰老的起始和推进受严密调控网络的精确控制。随着生长发育,玉米植株下部老叶不断地变黄、干枯,而新叶不断从茎尖伸展出来,营养生长期衰老叶片中的营养物质被再利用供给新叶生长,生殖生长期衰老叶片中的营养物质被转运到发育的籽粒中储存。延缓叶片衰老可以提高玉米的生物产量、籽粒产量和氮利用率。因此,玉米叶片衰老机制的研究可为叶片功能期的遗传改良奠定基础。与拟南芥相比,玉米叶片衰老调控机制的研究报道还较少。相关研究表明,糖分转运、源-库互作和活性氧等是生理水平上影响玉米叶片衰老的重要因素[75-76],阻止玉米授粉会导致源-库互作和糖分转运方式发生转变,进而诱导叶片早衰的起始,其中糖分转运相关基因的表达模式与拟南芥中衰老过程中基因表达模式相似[77-78]。
随着高通量测序的发展,研究者利用不同的组学分析,鉴定到一定数量叶片衰老调控相关的蛋白质、基因和miRNAs,但其功能和相关的分子机制尚不清楚。利用GWAS和转录组整合分析,SEKHON等[79]鉴定到8个与玉米叶片衰老显著性相关的基因,其中包括海藻糖-6-磷酸合成酶、木聚糖合成酶和NAC转录因子编码基因,这些基因均被证明参与了模式植物叶片衰老的调控。利用早衰突变体的Small RNA-seq和转录组分析,CHAI等[80]和WU等[81]鉴定到9个衰老相关的miRNAs和数个关键衰老相关基因,miRNAs包括zma-miR156,zma-miR159,zma-miR167,zma-miR171,zma-miR172,zma-miR395,zma-miR399,zma-miR408和zma-miR529;关键衰老基因包括Zm00001d016441(tyrosine aminotransferase)、Zm00001d036370(chitinase)、Zm00001d024767(aspartic proteinase A1)、Zm00001d014993(luminal-binding protein 2)、Zm00001d049217(SAG12)、Zm00001d043025(WRKY33)、Zm00001d003412(P-type R2R3 Myb protein)、Zm00001d012427(ZAT11)等[80-81]。ZHANG等[82]通过转录组分析,鉴定到了4 552个叶片自然衰老相关的差异表达基因。迄今,在玉米上仅有ZmVQ52(VQ motif containing protein Gene 52)、ZmSnRK1s、NAC7、knotted1和les1等叶片衰老相关的基因或突变体研究报道,其中叶片衰老正调控基因ZmVQ52和ZmSnRK1s的功能在拟南芥上进行了转化验证[38,83],而NAC7正向调控叶片衰老的功能在玉米中经过了转基因验证[84]。此外,叶片衰老负调控基因knotted1的功能在烟草转基因材料中得到了验证[85],司伟娜等[86]仅对les1进行了初步定位。综上所述,尽管前人对玉米叶片衰老的机制开展了一些研究,但对其代谢调控的分子机制仍知之甚少,极大地限制了玉米相关农艺性状的遗传改良。
3 RNA解旋酶基因在玉米熟期遗传改良中的应用
随着社会经济的发展和农业生产模式的转变,黄淮海夏玉米生产迫切需要实现籽粒机械化收获,而玉米生产中适于籽粒机械化收获的品种较少,急需玉米种质资源和育种技术的革新。高产、稳产、宜机收是当前玉米育种的主要目标,优良玉米新品种应该具有抗逆、抗病、叶片功能期长、籽粒灌浆速率快、籽粒脱水速率快、站秆性好等特点,叶片的适时衰老是制约育种目标实现的关键因素之一。玉米叶片衰老起始调控相关的RNA解旋酶基因通过叶片功能期适中有利单倍型的筛选,应用于成熟期植株衰老和籽粒脱水快的种质材料,进而选育适宜籽粒机收的玉米新品种。
随着人们对肉制品需求的不断增长,对作为饲料的玉米需求量和营养品质提出了更高的要求。青贮玉米不仅具有较高的生物产量,而且提供牲畜所需要的蛋白质,因而越来越受到重视。优良的青贮玉米品种应具有抗病、生物产量高、整株粗蛋白含量高、中性洗涤纤维含量低、植株淀粉含量高、持绿性好的特点。叶片持绿期长可为获得高生物产量和淀粉含量提供保障。RNA解旋酶主要参与植物环境响应调控,其功能丧失往往导致植株的早衰,非生物抗性和抗病性的衰退,而RNA解旋酶功能的增强和维持能够促进玉米持绿性提高,可应用于青贮玉米持绿性的改良。
4 展望
植物叶片衰老受控于复杂的调控机制,仅有的少数研究表明,RNA解旋酶在调控叶片衰老过程起始中发挥着极其重要的作用。虽然玉米等作物中RNA解旋酶的研究较少,但随着研究的深入,相关突变体和基因将被不断发掘,将为相关农艺性状的遗传改良提供必要的基因资源和理论依据。玉米宜机收新品种选育时既要注重叶片功能期的适当延长,又要在成熟期叶片快速衰老,促进籽粒脱水以达到籽粒机收的要求,精确控制叶片衰老的起始和进程是解决该问题的关键。因此,模式植物中RNA解旋酶功能研究不仅拓宽了人们对于植物叶片衰老调控机制的理解,而且为玉米机收品种的选育提供了一条崭新的途径。RNA解旋酶可能通过无义介导的 mRNA降解、端粒酶活性、Per-mRNA剪接、mRNA代谢等途径参与非生物逆境响应的调控,相关研究对于玉米抗逆性遗传改良具有重要的意义。玉米生长过程中,面临着大风、高温、干旱、涝渍等自然灾害,这些不利因素会引起植株的早衰和死亡,往往造成巨大的产量损失。对玉米RNA解旋酶参与非生物逆境响应机制进行研究,有助于通过遗传改良达到抗逆性与叶片功能期的优化,进而达到增产的目的。
虽然,模式植物RNA解旋酶的研究进展可为玉米的遗传改良奠定一定的理论基础,但仍存在若干瓶颈问题亟待解决。首先,植物叶片衰老是一个复杂的生物学过程,衰老起始和发展精确调控的分子机制还不清楚,亟待关键基因的分离和功能验证;其次,RNA解旋酶通常具有多种生物学功能,控制不同生物途径和农艺性状,作物遗传改良中难以达到平衡和优化;再次,玉米RNA解旋酶的研究较少,尚不能满足遗传改良的需求,需要开展相关研究,鉴定关键RNA解旋酶的功能和解析相关的分子机制;最后,夏玉米生产中不同气象灾害频发,通常造成严重的产量损失,农艺性状遗传改良需要多基因聚合,多种性状协调和平衡。随着玉米相关研究的开展,应用策略的革新,这些问题将会逐步得到解决。
猜你喜欢
华北农学报(2022年3期)2022-07-11
中国农学通报(2022年13期)2022-05-31
理财周刊(2022年4期)2022-04-30
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年1期)2022-02-28
作文·小学低年级(2021年10期)2021-01-25
学苑创造·A版(2020年12期)2020-01-07
科学种养(2017年6期)2017-06-13
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
