
新疆黑蜂基因组选择信号分析
2022-05-31 08:33:30何金明孙智禹刘玉玲牛庆生
中国畜牧兽医 2022年5期
何金明,孙智禹,刘玉玲,徐 凯,牛庆生
(吉林省养蜂科学研究所,吉林 132108)
新疆黑蜂是一个被列入中国《中国畜禽遗传资源志·蜜蜂志》的本地西方蜜蜂地方品种,中心产区为阿尔泰山、天山山脉及伊犁河谷地区,是新疆地区特有的、与当地气候环境及蜜源等生态条件相适应的优良蜂种,特别适合新疆北疆浅山区丘陵地带饲养,具有抗寒、抗逆、抗病、产卵力强、采集力强、节省饲料及蜂蜜纯天然且含天然药用成分等优点[1]。近年来,随着饲养方式的改变和引入蜂种的大量应用,新疆黑蜂的饲养范围逐年缩小,饲养量也不断减少。然而,饲养过程中的盲目引种导致新疆黑蜂杂化程度非常严重,纯种新疆黑蜂数量急剧下降。2006年,新疆黑蜂被列为《国家级畜禽遗传资源保护名录》的濒危物种之一,对新疆黑蜂的保护工作已迫在眉睫[2-3]。
中国西方蜜蜂(Apismellifera)遗传资源较为丰富,既有新疆黑蜂、东北黑蜂及浙江浆蜂等地方特色品种,又有意大利蜜蜂、卡尼鄂拉蜂等引入蜂种,不同蜂种在外部形态、生产性能、抗病力及适应性等方面具有较大差异,与各蜂种在进化过程中受到的长期自然选择和人工定向选择密切相关。选择作用会使生物群体在世代传递过程中的某些基因型个体比例发生变化,继而在基因组上留下相应的选择印迹,即选择信号(selection signal)[4]。利用基因组上的选择信号进行群体遗传学分析有助于了解种群的进化历史,并对研究物种的种质特性、遗传育种等问题提供帮助。随着基因组学和高通量测序技术的快速发展,大范围的基因组选择信号得以快速检测,为畜禽种质资源的鉴定和保护利用提供了新的方法[5]。通过全基因组重测序技术检测选择信号以挖掘动物优良性状相关候选基因已被成功应用于牛[6-7]、猪[8]、鸡[9]、羊[10-11]等畜禽中,并鉴定出了多个与繁殖、毛色、生长等经济性状相关的重要基因。动物基因组选择信号分析的成功应用为揭示蜜蜂遗传资源进化历史及发掘其优良性状相关潜在遗传学基础提供了新的研究方向。当前关于新疆黑蜂的多数研究仅限于常规生产性能、繁殖性能、蜂蜜王浆品质的测定与比较,对其种质资源的鉴别仅能根据形态特征等传统方法进行判断,缺乏对其品种优良特性及候选基因的研究,其遗传进化关系尚不明确[12-13]。鉴于上述情形,本研究利用全基因组重测序技术对新疆黑蜂和其他引入品种进行不同蜂种间的遗传差异检测,分析新疆黑蜂的遗传进化关系,并通过选择信号分析挖掘新疆黑蜂特异性状的相关基因,以期为制定更有效的保种措施和进一步的蜂种资源利用与开发提供理论依据。
1 材料与方法
1.1 材料
供试蜂种为来源于吉林省养蜂科学研究所(国家级蜜蜂基因库)的新疆黑蜂(Xinjiang Black bee,XJB)、意大利蜜蜂(Apismelliferaligustica,ITA)、欧洲黑蜂(Apismelliferamellifera,EUR)、卡尼鄂拉蜂(Apismelliferacarnica,CAR)、高加索蜂(Apismelliferacaucasica,CAU)的核心保种群。于2020年5月15日从各蜂种中分别选取10群培育蜂王,2020年6月1日从每群中分别选取1只刚羽化出房的处女蜂王进行试验。长白山中华蜜蜂(Apismelliferacaucasica,A.c.c)采自位于吉林省敦化市的长白山中华蜜蜂保种场,选取1只未交尾的处女蜂王作为群体遗传分析的参考群体。收集的蜂王样本立即投入液氮中,返回实验室后(约4 h)立即放入-80 ℃保存,用于基因组DNA提取。
1.2 方法
1.2.1 基因组重测序数据获取 使用天根生化科技(北京)有限公司组织基因组DNA试剂盒提取蜜蜂组织基因组DNA,构建500 bp双末端测序文库,利用Illumina HiSeq PE150平台进行测序。
1.2.2 测序数据质控、过滤和处理 为了得到有效且高质量测序数据(clean reads),需要对Illumina HiSeq PE150平台获得的原始测序数据(raw reads)进行严格过滤。过滤原则如下:①去除含有接头序列的reads;②去除单端测序reads中含N比例>10%的paired reads;③仅保留单端测序reads中含低质量碱基数<50%的序列。将质控后的高质量数据通过BWA(参数:mem-t-k32-M-R)软件与西方蜜蜂参考基因组(https:∥ftp.ncbi.nlm.nih.gov/genomes/all/GCF/003/254/395/GCF_003254395.2_Amel_HAv3.1/GCF_003254395.2_Amel_HAv3.1_genomic.fna.gz)进行比对,获得比对参考基因组后的BAM文件,用SAMTOOLS对BAM文件进行排序、去除重复,并进行SNP和Indel检测,获得高质量的遗传变异位点。使用ANNOVAR软件对识别出的高质量遗传变异进行SNP信息注释。
1.2.3 生物信息学分析 基于全基因组重测序技术检测出的高质量SNP,采用Mega 6.0在线软件(http:∥www.megasoftware.net/)的邻接法(Neighbor-Joining)构建系统进化树;使用Plink 1.09在线软件(https:∥www.cog-genomics.org/plink/1.9/)进行主成分分析(principal component analysis,PCA);利用Admixture 3.0在线软件(http:∥software.genetics.ucla.edu/admixture/)进行群体遗传结构分析;应用Haploview 4.1在线软件(http:∥www.broad.mit.edu/mpg/haploview/)进行连锁不平衡分析。 通过Plink 1.09软件,基于群体遗传分化指数(Fst)和核苷酸多样性比值(θπ)两种方法筛选排名5%的候选选择区域,进行选择信号的筛选。利用David在线软件(https:∥david.ncifcrf.gov/)对提取到的受选择区域候选基因进行GO功能和KEGG通路富集分析。
2 结 果
2.1 新疆黑蜂基因组SNP鉴定
新疆黑蜂全基因组重测序的数据经质控后,最终鉴定出了1 728 216个SNPs,新疆黑蜂中有73 812个SNPs位点位于编码区,占4.3%;有1 159 436个SNPs位点位于内含子区,占67%;394 942个SNPs位点位于基因间区,占22.85%。新疆黑蜂基因组中存在18 526个非同义突变位点,可引起氨基酸的改变,可作为该蜂种遗传特性候选SNP位点。
2.2 系统进化树分析
通过对新疆黑蜂、意大利蜜蜂、高加索蜂、欧洲黑蜂、卡尼鄂拉蜂的SNP进行系统发育关系分析,结果见图1。由图1可知,意大利蜜蜂先与欧洲黑蜂聚在一支,其次是卡尼鄂拉蜂、新疆黑蜂,最后是高加索蜂,与其他蜂种相比,新疆黑蜂与高加索蜂亲缘关系较远;新疆黑蜂所有个体聚为一小支,其群体血统单一,与其他蜂种无个体交叉现象。
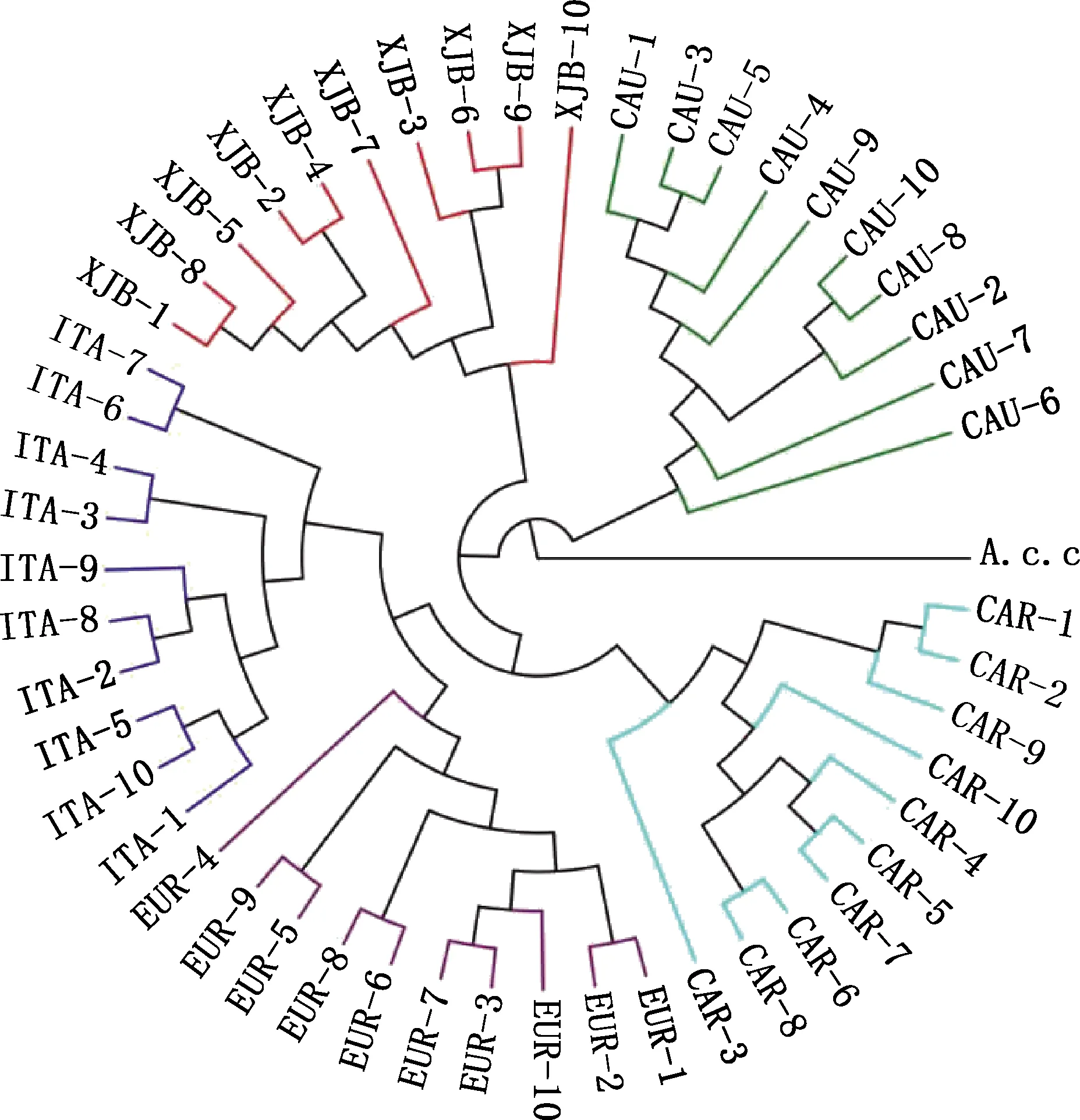
XJB,新疆黑蜂;ITA,意大利蜜蜂;CAU,高加索蜂;EUR,欧洲黑蜂;CAR,卡尼鄂拉蜂;A.c.c,中华蜜蜂。下同XJB,Xinjiang Black bee;ITA,Apis mellifera ligustica;CAU,Apis mellifera caucasica;EUR,Apis mellifera mellifera;CAR,Apis mellifera carnica;A.c.c,Apis cerana cerana.The same as below图1 新疆黑蜂及4个引入蜂种的系统进化树Fig.1 Phylogenetic tree of Xinjiang Black bee and 4 introduced bees
2.3 SNP主成分分析
对所有试验个体使用最大的2个主成分,第一主成分(PC1)和第二主成分(PC2)作图(图2)。由图2可知,第一主成分将高加索蜂(CAU)分开, 第二主成分将其余蜂种分成了新疆黑蜂(XJB)、意大利蜜蜂(ITA)以及欧洲黑蜂(EUR)和卡尼鄂拉蜂(CAR)聚为一簇的群体,新疆黑蜂与其他蜂种之间存在一定的差异。

图中每一个点代表一个样品,不同的颜色代表不同的蜂种Each point represents a sample,different colors represent different bees图2 主成分分析结果Fig.2 Results of principal component analysis
2.4 群体遗传结构分析
通过对5个蜂种的SNP进行群体遗传结构分析,绘制出了Structure推叠图(图3)。由图3可知,假设祖先群体亚群数K=2时,50个个体被划分为了2个亚群体(深蓝和浅蓝),高加索蜂、欧洲黑蜂、意大利蜂血统比较纯,新疆黑蜂和卡尼鄂拉蜂的个体有2种组成成分,说明其很有可能是由2个祖先亚群杂交而来;K=3时,其他蜂种没有明显变化,意大利蜂分离出来;当K>3时,新疆黑蜂被完全分化出来,其所属祖先亚群与意大利蜂、卡尼鄂拉蜂、高加索蜂、欧洲黑蜂完全不同。
2.5 连锁不平衡分析
由图4可知,各蜂种的衰减速度是欧洲黑蜂>卡尼鄂拉蜂>高加索蜂>新疆黑蜂>意大利蜜蜂,即意大利蜜蜂的连锁不平衡衰减速度最小,其次是新疆黑蜂,表明在进化过程中这2个群体与其他群体相比受驯化程度较高,受选择强度较大。

图中分别展示了预设为2~6个群体的结构;每份样品用一个垂直线条表示,每个垂直线条的颜色表示该样品祖先种群的比例The figure shows the structure of the predetermined 2 to 6 groups,respectively;Each sample is respresented by a vertical line,and line color indicates ancestral population proportion图3 群体遗传结构分析结果Fig.3 Results of population genetic structure analysis
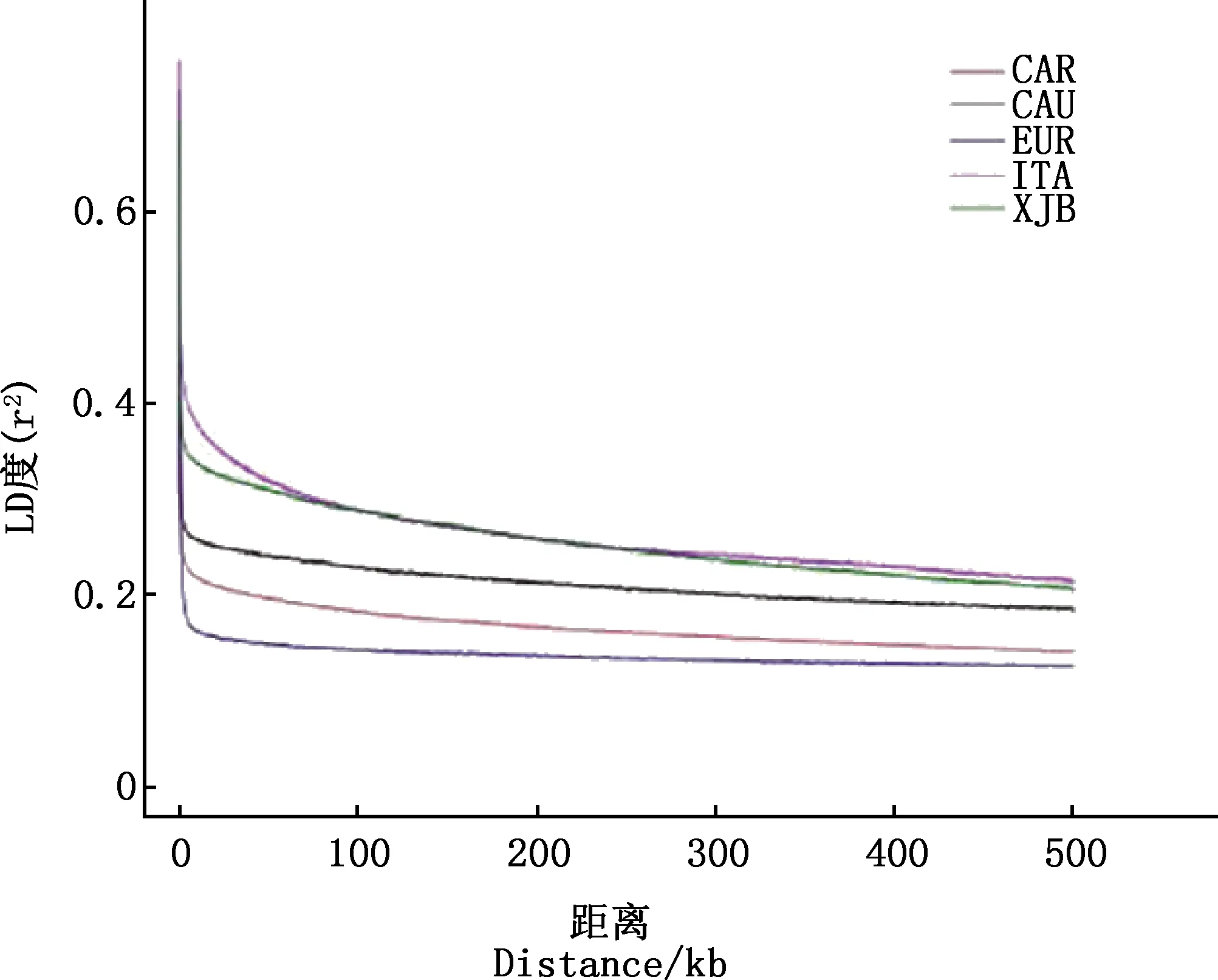
图4 连锁不平衡衰减图Fig.4 Linkage disequilibrium patterns map
2.6 选择信号分析
通过计算基因组的θπ和Fst值,对筛选出的前5%区域的受选择信号进行分析,将受选择的基因进行GO功能富集分析,结果显示,新疆黑蜂受选择基因主要富集在代谢、繁殖、信号转导等生物过程,细胞组分主要富集在细胞器、核小体、染色质和膜等,分子功能主要富集在有机环状化合物结合、离子结合等功能,其中在这些主要富集的通路中共筛选出了细胞色素P450 314A亚家族多肽1(cytochrome P450 family 314 subfamily A polypeptide 1,Cyp314A1)、毛透明蛋白(Trichohyalin-like)、卷曲螺旋结构域蛋白112(coiled-coil domain-containing protein 112,CCDC112)、山梨醇脱氢酶(Sorbitol dehydrogenase)、丝氨酸蛋白酶抑制剂3(serine protease inhibitor 3,SPI-3)、铁蛋白(soma ferritin,Fer3HCH)、张力蛋白1(tensin-1,TNS1)、磷酸酶与张力蛋白同源物(phosphatase and tensin-like,Pten)、腺苷琥珀酸裂解酶(adenylosuccinate lyase,ADSL)、章鱼胺受体β2R(octopamine receptor beta-2R,Octβ2R)、G补缀FHA域血管生成因子1(angiogenic factor with G patch and FHA domains 1,AFFG1)、鸟嘌呤核苷酸交换因子17(rho guanine nucleotide exchange factor 17,ARHGEF17) 12个候选基因(表1),这些基因可能与新疆黑蜂的抗寒、产卵力强、抗病等生物学特性相关,参与了机体的繁殖、生长发育及免疫等生理活动。KEGG通路富集分析结果表明,卡尼鄂拉蜂与新疆黑蜂基因组相比,新疆黑蜂受选择基因主要富集于嘌呤代谢、胞吞作用等信号通路;高加索蜂与新疆黑蜂基因组相比,新疆黑蜂受选择基因主要富集于胞吞作用、磷酸肌醇代谢等信号通路;欧洲黑蜂与新疆黑蜂基因组相比,新疆黑蜂受选择基因主要富集于物质代谢等信号通路,意大利蜂与新疆黑蜂基因组相比,新疆黑蜂受选择基因主要富集于嘌呤代谢等信号通路(图5)。
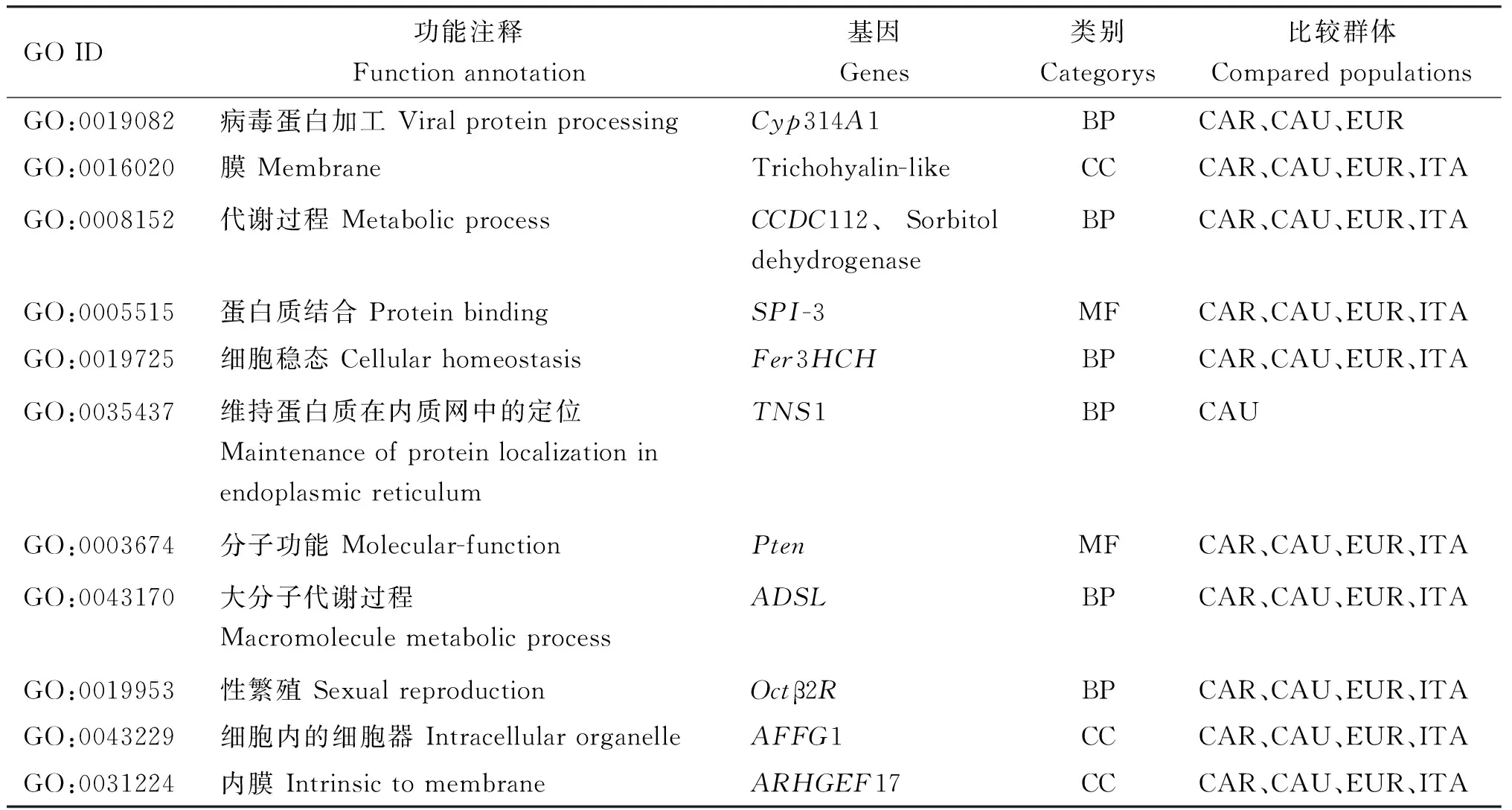
表1 新疆黑蜂受选择基因GO功能富集分析

A~D,分别为与卡尼鄂拉蜂、高加索蜂、欧洲黑蜂、意大利蜂相比新疆黑蜂受选择的KEGG通路富集分析图A-D,KEGG pathway enrichment analysis of selected genes in Xinjiang Black bees compared with Apis mellifera carnica,Apis mellifera caucasica,Apis mellifera mellifera,Apis mellifera ligustica图5 新疆黑蜂受选择基因KEGG通路富集分析Fig.5 KEGG pathway enrichment analysis of selected genes in Xinjiang Black bee
3 讨 论
据报道,俄国人于1900年将黑色蜂种带入了中国新疆伊犁和阿勒泰地区,1919年又经哈萨克斯坦将黑蜂带入了新疆布尔津县,1925-1926年又将黑蜂带到了新疆伊宁饲养,慢慢发展到了整个伊犁地区,经过百余年的长期混养、自然杂交和人为选育逐渐形成了适应新疆气候环境的蜂蜜高产型地方品种[1]。但是关于该蜂种是何起源未见详细记录。一直以来,新疆黑蜂被认为是欧洲黑蜂或高加索蜂的后代[14-15],但都没有相关的遗传学证据。本研究对国家级蜜蜂基因库(吉林)异地保存的新疆黑蜂保种群体进行遗传学分析,发现新疆黑蜂与卡尼鄂拉蜂、意大利蜂、欧洲黑蜂亲缘关系较近,而与高加索蜂的亲缘关系较远,所以暂不支持新疆黑蜂来源于高加索蜂。西方蜜蜂的起源进化和分类问题一直存在争议,在目前已被确认的29个西方蜜蜂亚种中,根据形态学及线粒体分子数据被划分为4个主要谱系:A系(非洲)、M系(西欧)、C系(东欧)、O系(中东)[16]。本研究系统进化树结果与上述分类略有不同,可能是由于分析所用的数据集和具体的分析方法不同。此外,结合系统发育分析、主成分分析及群体遗传结构和连锁不平衡分析结果表明,新疆黑蜂在经历了长时间的人工选择及对中国新疆地区的气候环境产生高度适应性后,与其他引入蜂种已经产生了明显的遗传差异,形成了独立的遗传背景,且在进化过程中受到的选择强度较大,推测可能是由于长期的自然选择和人工选育造成的。
新疆黑蜂对中国新疆地区的气候、蜜源等生态条件逐渐形成了很强的适应性。本研究筛选出了毛透明蛋白(Trichohyalin-like)、山梨醇脱氢酶(Sorbitol dehydrogenase)等新疆黑蜂受选择的候选基因,这些基因已被证明与昆虫的抗寒及能量代谢有关。毛透明蛋白是一种富含谷氨酸、丙氨酸、精氨酸、赖氨酸、丝氨酸的结构蛋白,其特征氨基酸也是抗寒物质中的关键氨基酸。研究显示,不同温度梯度下的烟粉虱MED隐种和Asia Ⅱ 3隐种Trichohyalin基因的表达存在差异,且该基因在抵御低温过程中发挥了一定的调控作用[17],有助于烟粉虱在低温条件下生存;山梨醇脱氢酶对昆虫滞育期能量代谢的调控起到重要作用,它可以将越冬期的抗寒物质山梨醇转化成糖原,进而分解成可供昆虫生长发育的能源物质[18]。新疆黑蜂可以在恶劣的气候、地理条件下生存,是中国唯一能够在野外生存的西方蜜蜂类型,而这些候选基因的功能将有助于新疆黑蜂抗寒、越冬性能强等生物学特性的形成。
本研究筛选出的CYP314A1、CCDC112、Octβ2R等新疆黑蜂受选择的候选基因在昆虫以及人类繁殖、产卵等生命活动中发挥着重要作用。研究发现,CYP314A1基因属于CYP314家族成员,与昆虫中诱导卵子发生和抗药性形成等生理活动密切相关[19-20];CCDC112是与高龄妇女产卵有关的差异蛋白之一[21];Octβ2R基因可以激活多种信号通路以诱导产卵,而Octβ2R基因纯合突变则导致果蝇雌性个体不育,它们虽表现出正常的求爱、交配、精子储存和交配后排斥行为,但最后无法产卵[22]。新疆黑蜂体内CYP314A1、CCDC112、Octβ2R等繁殖候选基因在进化过程中受到选择可能有利于该蜂种高繁殖能力的形成,因而具有产卵力较强、子脾面积大且春季繁殖快等特点,这些基因可以作为日后研究新疆黑蜂优异繁殖性能的候选基因。此外,本研究中还筛选出了一些与生长发育相关的基因,它们也在新疆黑蜂进化的过程中受到了选择,如与细胞凋亡、增殖及血管生成有关的Kruppel样因子6(Kruppel-like factor 6,KLF6)[23];通过诱导细胞凋亡来参与抵抗家蚕核型多角体病毒侵染的细胞色素C(Cytc)[24];在细胞黏附、运动、生长等细胞生物学活动中发挥着重要作用的TNS1基因[25];协作参与维持基因组稳定性、修复DNA、参与控制基因重组的Pten基因[26];与肌肉、生长、维持细胞代谢有关的ADSL基因[27];高表达于血管内皮细胞,编码蛋白具有强力血管生成作用的AFFG1以及对维持内皮间连接起着非常重要作用的ARHGEF17等基因[28-29]。KLF6、TNS1等上述基因的表达调控可能在新疆黑蜂的生长发育过程中起到了重要作用,使其更能应对伊犁北疆寒冷多风的恶劣生存环境。
新疆黑蜂在抗病力方面也优于其他西方蜜蜂品种,本研究筛选出了SPI-3、Fer3HCH等多个与昆虫免疫相关的基因。已有研究发现,丝氨酸蛋白酶抑制剂(Serpins)是昆虫体内一类非常重要的蛋白酶抑制剂,昆虫免疫应答的重要组成部分,其家族成员SPI-1和SPI-2基因在正常大鼠肝脏中表达,但SPI-3基因在正常大鼠中几乎处于沉默,却在大鼠肝脏急性炎症期间以及大鼠脑短暂缺血后变得短暂活跃,表明SPI-3基因可能参与了针对炎症应激的细胞防御反应[30-31]。昆虫中的Fer3HCH基因通常存在于液泡系统和血淋巴中,研究认为该基因是赤拟谷盗的一种潜在的抗菌效应因子,在抵御病原感染方面具有一定作用;另外,果蝇体内基因过表达试验结果表明,Fer3HCH基因在保护机体免受氧化应激胁迫等方面发挥着重要作用[32]。以上与免疫相关的候选基因在新疆黑蜂体内受选择,有利于该蜂种形成免疫力强的体质,从而具有较强的抗病力。
4 结 论
本研究通过全基因组重测序技术在新疆黑蜂中共鉴定出1 728 216个SNPs,其中18 526个非同义突变位点是后续研究新疆黑蜂遗传特性候选的SNPs位点;筛选到了CYP314A1、Trichohyalin-like、CCDC112、Sorbitol dehydrogenase、SPI-3、Fer3HCH、TNS1、Pten、ADSL、Octβ2R、AFFG1、ARHGEF17共12个候选基因,解析了新疆黑蜂的抗寒、产卵力强、抗病等优异性状形成的分子机理,为新疆黑蜂种质资源评价、保护及开展分子标记辅助选择提供了理论依据。
猜你喜欢
青年生活(2019年5期)2019-09-10 07:22:44
中国蜂业(2018年7期)2018-07-27 03:50:00
——黑蜂的保护利用建议
中国蜂业(2018年1期)2018-01-21 03:25:26
——新疆黑蜂素材后代初步鉴定检测结果
中国蜂业(2017年12期)2018-01-05 05:45:10
——新疆黑蜂素材后代初步鉴定检测结果
中国蜂业(2017年11期)2017-12-29 06:32:33
小天使·一年级语数英综合(2017年9期)2017-10-20 21:12:02
安徽农业科学(2017年12期)2017-05-30 06:12:26
——沿着历史上分布的踪迹追溯黑蜂信息
中国蜂业(2017年5期)2017-01-15 11:49:27
阅读与作文(小学高年级版)(2016年5期)2016-05-10 23:32:23
阅读与作文(小学低年级版)(2016年1期)2016-03-12 22:20:48
