
不同生长年限滇重楼的生物量分配与异速生长研究
2022-05-31 11:49王元忠张金渝
中草药 2022年11期
王元忠,张 霁,沈 涛,张金渝*
不同生长年限滇重楼的生物量分配与异速生长研究
王元忠1,张 霁1,沈 涛2,张金渝1*
1. 云南省农业科学院药用植物研究所,云南 昆明 650200 2. 玉溪师范学院化学生物与环境学院,云南 玉溪 653100
比较不同产地和生长年限栽培滇重楼var.生物量和性状差异,并对植株生物量积累和生长发育规律进行异速生长分析。使用Pearson相关分析研究滇重楼根茎生物量与采集地经纬度、气候因子间的相关性;单因素方差分析(One-Way ANOVA)与独立样本T检验用于样品间植物性状的差异显著性比较;利用标准化主轴回归分析(standardized major axis regression)对滇重楼的生物量积累和生长发育规律进行异速生长研究。研究表明12个不同产地的5年生滇重楼根茎生物量平均值在1.19~19.73 g,不同产地之间具有显著差异(<0.05),但该差异与产地经纬度、年均温度、年均降水量之间不存在显著相关性(>0.05)。滇重楼植株的根茎生物量均随生长年限增加而增加,种植5年后根茎生物量一直呈现快速积累趋势。相同生长年限植株中,云南省元阳县样品根茎生物量显著高于姚安县样品(<0.05),并且数值具有更高的变化范围。滇重楼植株的总生物量、株高、叶生物量、叶长、叶宽、叶面积均随生长年限增加而增加,比叶面积均随生长年限增加而减小。云南省元阳县滇重楼样品根茎生物量占总生物量的比例随生长年限增加而增加,云南省姚安县样品根茎生物量比例不随生长年限变化而变化。滇重楼植株总生物量和根茎生物量、株高、叶生物量、比叶面积之间均存在异速生长关系。与云南省姚安县滇重楼样品相比,云南省元阳县滇重楼样品根茎生物量积累速率高、株高较矮、叶生物量积累速率较低、比叶面积较大。培育高产滇重楼,应考虑选择株高较矮、叶片较大而薄的地理种源;人工栽培滇重楼时需要注意合理控制光照等因素,从而提高滇重楼药用部位产量。
滇重楼;生物量;异速生长;功能性状;产量;生长
植物可利用的资源总量往往是有限的,其对某些功能性状资源投入的增加,必然意味着对其他性状资源投入的减少。也就是说,在有限的资源环境中,植物会根据所处生境的资源条件在功能性状之间进行资源优化配置,通过调整自身功能,实现生存、生长以及繁殖的需要,最终表现在植物器官的结构、生理及化学性状方面[1]。例如,比叶面积(specific leaf area)是单位干物质的叶面积,反映了叶片捕获光照资源的能力,与相对生长速率和净光合速率成正比。比叶面积低意味着叶片大而薄,植株可以更好地截获光合有效辐射[2],进而影响植株生物量的积累和分配[3]。
异速生长(allometry)是有机体某一部位或某一指标的不等速变化,即生物体各部分的生物量、形态特征等数值的变化与生物个体大小(通常以总生物量来表示)之间存在相对变化关系[4]。植物的异速生长被视为对众多选择压力和胁迫所作出的响应[5]。开展药用植物的异速生长研究,可以用于分析药用植物生长发育及品质的形成规律与环境之间的关系,为药用植物资源评价研究提供生态学的思路[6-9]。
滇重楼var.(Franch.) Hand. -Mazz.为重楼属多年生草本植物,是云南特色药用植物[10-11]。前期研究发现重楼属植物的形态多样性受到遗传与地理环境的影响[12]。进一步研究显示,产地纬度、温度、降水量等因素影响着重楼叶片的光合过程与糖代谢,进而导致不同种源药用部位的产量差异[13]。因此,围绕影响重楼根茎生物量的光合部位发育情况,从光合产物分配的角度,探讨环境因子对滇重楼药用部位产量的影响具有重要意义[14-15]。
本研究通过测定不同居群滇重楼生物量及形态结构指标,比较根茎生物量和与之相关的地上部分形态指标的产地差异性,以及随生长年限增长的动态变化规律,为重楼优良种源筛选及优化人工栽培管理提供参考数据。
1 材料与方法
1.1 材料
于2017年9~10月,在云南省、四川省、陕西省12个地点采集不同年限栽培滇重楼样品(表1)。采样地年均温和年均降水量信息从Worldclim数据库(http://www.worldclim.org/)提取。样品经云南省农业科学院药用植物研究所张金渝研究员鉴定为滇重楼var.(Franch.) Hand. -Mazz.。其中红河州元阳县小新街乡滇重楼为3~8年生,楚雄州姚安县前场镇滇重楼为3~9年生。每生长年限均收集9~16株完整个体。用卷尺测量株高,统计叶片数,并使用直尺测量每株植物每片叶片的叶长、叶宽。植株带回实验室,烘干,测定不同部位生物量。其余10个地点滇重楼均为5年生,取完整根茎带回实验室,烘干称定质量。
表1 滇重楼采样信息
Table 1 Sampling information of var.
居群采样地点纬度(N)经度(E)年均温度/℃年均降水量/mm 1云南省元阳县23.00°103.02°16.21336 2云南省姚安县25.52°101.38°14.3 797 3云南省玉溪市24.43°102.44°15.1 934 4云南省丽江市26.88°100.19°12.9 787 5云南省文山市23.24°104.07°14.91523 6云南省昆明市25.16°103.01°14.5 928 7云南省保山市24.57° 98.49°14.61410 8四川省彭州市31.18°103.81°15.2 958 9云南省巍山县25.31°100.32°14.4 861 10陕西省勉县33.51°106.64° 9.7 771 11云南省云县24.44°100.12°20.81130 12云南省景谷县23.41°100.40°20.61375
1.2 性状测定与统计分析
根据叶片测量结果,计算叶长及叶宽平均值,并由公式(1)、(2)计算叶面积、比叶面积。
叶面积=叶长×叶宽×叶面积系数 (1)
比叶面积=叶面积/叶片生物量 (2)
公式(1)中滇重楼叶面积系数根据文献报道的滇重楼叶长、叶宽、叶面积数据,确定为0.729 5[16-17]。
各数据经对数转换(log),使用SPSS 22.0统计分析软件中的Pearson相关分析法分析12个不同地点5年生滇重楼根茎生物量与采集地经纬度、年均温、年均降水量之间的相关性;采用单因素方差分析法(One-Way ANOVA)和Tukey多重比较法分别比较12个不同地点5年生滇重楼根茎生物量差异以及2个不同地点滇重楼生物量及地上部分性状随生长年限变化的差异;采用独立样本T检验比较同一年限2个不同地点滇重楼生物量及地上部分性状差异。另外,使用R 3.4 的软件包“smatr”中的标准化主轴回归分析(Standardized Major Axis Regression)功能,利用线性回归模型,对滇重楼的生物量积累和生长发育规律进行异速生长分析[18]:
log=log(3)
log是滇重楼单株根茎生物量、株高、叶生物量或比叶面积的对数,log是单株总生物量的对数,是线性回归关系的斜率,是线性回归关系的截距
2 结果与分析
2.1 相同生长年限不同产地滇重楼根茎生物量差异
单因素方差分析发现(表2),12个不同产地的5年生滇重楼根茎生物量具有显著差异(<0.05)。采自云南省保山市龙陵县河头乡样品根茎生物量达到19.73 g,显著高于其他产地样品;云南省玉溪市红塔区黄草坝村(13.30 g)和红河州元阳县小新街乡(9.96 g)样品根茎生物量次之。其余产地滇重楼样品根茎生物量为1.19~4.82 g,显著低于前3个产地。各产地中,云南省红河州元阳县小新街乡样品根茎生物量RSD最高,达到60.6%,云南省大理州巍山县三家村样品RSD最低,为28.5%。Pearson相关分析结果表明,滇重楼根茎生物量与其产地经纬度、年均温、年均降水量未表现出显著相关性。提示本研究中不同产地滇重楼根茎生物量之间存在的显著差异,可能与其他环境因素或遗传因素相关。
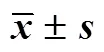
表2 不同产地5年生滇重楼根茎生物量比较(, n = 3)
不同小写字母差异显著(<0.05)
Different lowercase letters mean significant differences (<0.05)
2.2 滇重楼生物量及叶片性状随生长年限的动态变化
图1显示,2个产地滇重楼植株的根茎生物量均随生长年限增加而增加;相同生长年限植株中,元阳县样品根茎生物量显著高于姚安县样品(<0.05),并且数值具有更高的变化范围。不同居群滇重楼植株性状指标的进一步分析发现(表3),植株总生物量、叶生物量、叶长、叶宽、叶面积等性状虽在居群间呈现显著差异(<0.05),但各性状随植株随生长年限的增加,呈现相似的变化趋势。其中,生长8、9年的植株,其总生物量、株高、叶生物量、叶长、叶宽和叶面积显著高于生长3~4年的植株;而叶片数和比叶面积则显著低于3~4年生植株。大部分性状数值在植株生长至第6年或第7年时,与3~5年生的植株产生显著差异(<0.05),数值明显增加。比叶面积在居群间差异不显著,但植株生长至第5年或6年时,其数值明显降低(<0.05)。根茎生物量在总生物量中的占比受到不同种群和植株年龄的共同影响;元阳居群,7年和8年生植株,其根茎生物量与总生物量的比值显著高于其它植株(<0.05),而姚安居群,不同生长年限植株根茎生物量与总生物量的比值变化不显著(>0.05)。
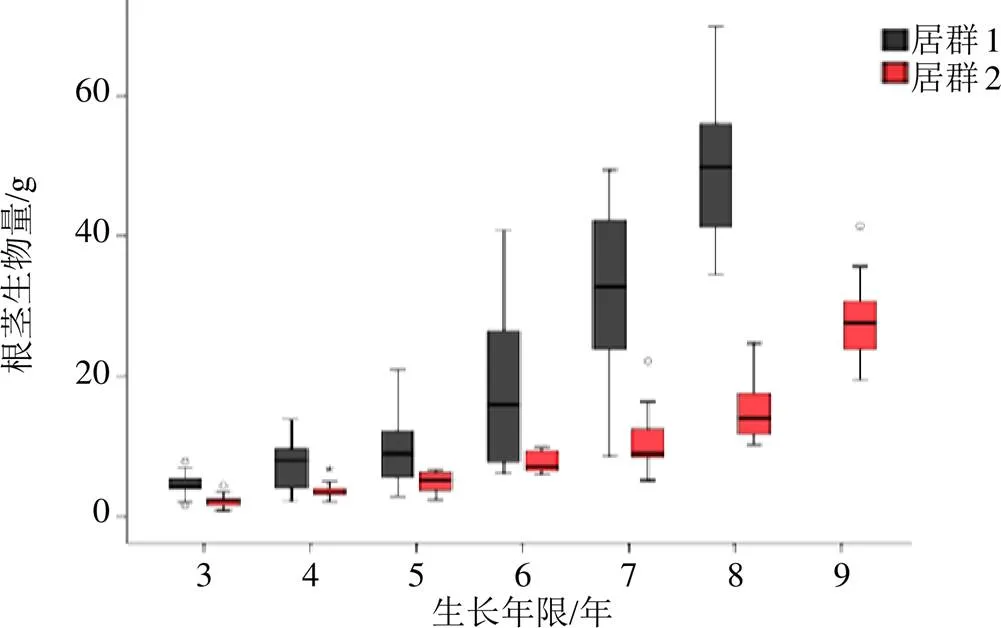
图1 不同产地滇重楼根茎生物量随生长年限的动态变化
表3 不同产地滇重楼植株性状指标动态变化
Table 3 Dynamic changes of traits of var. samples from different origins
居群编号生长年限/年总生物量/g株高/cm叶生物量/g叶片数叶长/cm 135.81±2.16 aB17.8±9.9 aA0.65±0.25 aB5±1 aA9.0±2.3 aB 49.61±3.97 aB20.5±9.8 aA0.85±0.31 abB6±1 abB10.1±2.1 abB 512.31±7.15 abB25.3±9.2 abA1.13±0.50 abB6±1 abA10.7±1.9 abB 622.40±15.28 bB28.3±15.3 abA1.73±1.16 bcB6±1 bA12.4±2.9 bcB 736.89±14.51 cB34.6±16.1 bA2.26±0.99 cdA7±1 bA14.4±3.3 cdB 855.91±13.72 dB36.0±8.7 bA3.05±1.52 dA7±1 bA15.9±2.4 dB 232.81±1.13 aA16.0±3.7 aA0.33±0.15 aA5±1 aA5.7±1.1 aA 44.72±1.22 abA22.3±6.5 abA0.56±0.15 abA5±1 aA7.1±1.3 abA 56.37±1.89 abA22.9±9.1 abA0.71±0.16 abA5±1 aA7.7±0.7 bcA 69.64±1.92 bcA28.1±10.0 abA0.99±0.24 bcA6±0 bA8.8±1.4 cA 714.44±6.12 cA30.5±12.0 bA1.55±0.52 cdA6±1 bA11.1±1.8 dA 820.51±5.21 dA46.5±15.0 cA2.17±1.11 dA7±1 bA12.4±1.3 deA 938.62±9.88 e67.6±16.0 d3.09±0.62 e7±2 b13.3±1.5 e 居群编号生长年限/年叶宽/cm叶面积/cm2比叶面积/(cm2 g−1)根茎生物量/总生物量 134.6±0.8 aB31.4±13.0 aB9.6±2.5 bA0.77±0.03 aA 44.6±0.8 aB31.4±13.0 aB9.6±2.5 bA0.77±0.03 aA 54.7±0.9 aB35.1±11.7 aB7.8±2.3 abA0.78±0.09 abA 65.2±1.2 aB41.8±14.4 aB7.5±3.7 abA0.80±0.06 abA 75.7±1.2 abA53.2±21.3 abB5.9±2.0 aA0.83±0.03 bcA 86.7±1.6 bB73.0±31.6 bcB5.2±1.2 aA0.87±0.02 cB 236.8±1.2 bB81.2±23.6 cB4.9±2.3 aA0.89±0.03 cB 43.0±0.4 aA12.6±3.9 aA10.1±5.4 cA0.78±0.05 aA 53.7±0.8 abA19.9±7.3 abA7.5±1.7 bcA0.77±0.06 aA 63.6±0.5 abA20.5±3.5 abA6.2±2.7 abA0.76±0.05 aA 74.8±1.2 bcA30.7±10.1 bcA5.3±2.3 abA0.78±0.03 aA 84.9±1.4 bcdA40.8±17.5 cdA4.4±1.7 abA0.78±0.03 aA 95.8±1.9 cdA54.0±24.1 deA3.9±1.0 aA0.73±0.07 aA
小写字母不同表示同一产地不同生长年限滇重楼样品数值之间有显著差异(<0.05);大写字母不同表示同一生长年限不同产地滇重楼样品数值之间有显著差异(<0.05)
Different lowercase letters mean significant differences between samples with different growing years (< 0.05); Different capital letters mean significant differences between samples collected from different origins with same growing years (< 0.05)
2.3 异速生长
由表4可知,2个产地的滇重楼植株总生物量和根茎生物量、株高、叶生物量、比叶面积之间均存在异速生长关系。图2显示,当总生物量较小时,元阳县滇重楼根茎生物量样品小于姚安县样品;但随着总生物量增大,元阳县滇重楼样品根茎生物量大于姚安县样品。叶片生物量变化规律与根茎生物量变化规律相反。当总生物量一定时,元阳县样品株高小于姚安县样品,但比叶面积大于姚安县样品。
表4 不同产地滇重楼异速生长方程
Table 4 Allometric equations of var.samples from different origins
异速关系(Y-X)居群方程R2P 根茎生物量-总生物量1log y=1.057 7 log x-0.155 30.994<0.001 2log y=0.983 1 log x-0.105 30.992<0.001 株高-总生物量1log y=0.537 1log x+0.747 50.517<0.001 2log y=0.600 6 log x+0.879 50.624<0.001 叶片生物量-总生物量1log y=0.776 1log x-0.833 10.839<0.001 2log y=0.902 3 log x-0.885 20.935<0.001 比叶面积-总生物量1log y=–0.444 0 log x+1.323 40.488<0.001 2log y=–0.553 7 log x+1.244 20.625<0.001
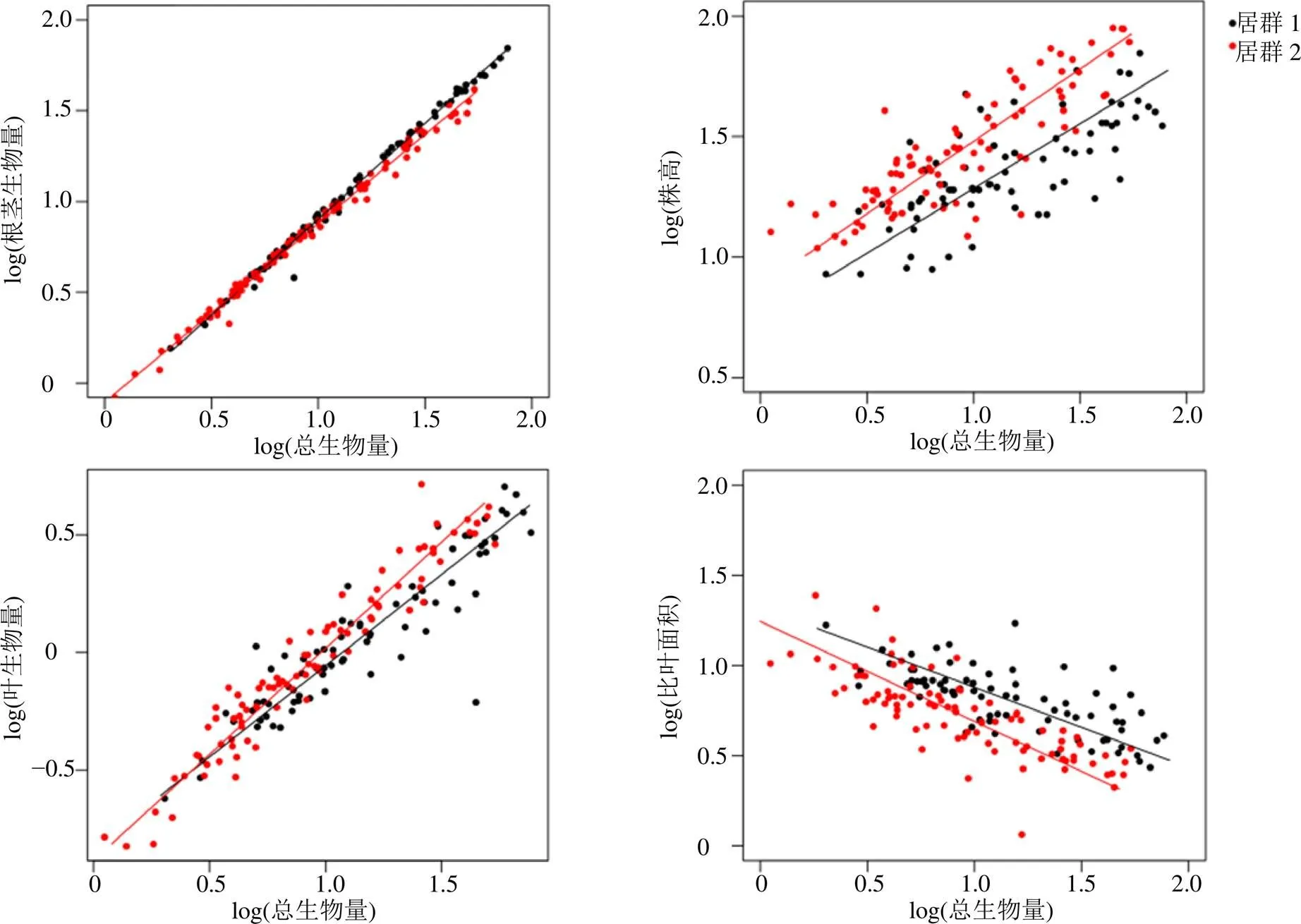
图2 不同产地滇重楼异速生长分析
3 讨论
云南属于低纬高原地区,立体气候明显,地形地貌、土壤类型复杂多样。同一年限不同产地滇重楼根茎生物量的差异可能受到环境因素影响。例如,温度对滇重楼的生长发育和光合速率有直接影响[19]。当温度在15~28 ℃时,滇重楼光合速率最高,有利于根茎生物量的积累;高于或低于此温度范围,光合速率均下降[20-22]。水分也是影响滇重楼生长发育的一个重要影响因素。每年11月中下旬,滇重楼地上部分倒苗,地下部分进入休眠期越冬。从倒苗到第2年3~4月出苗,这一时间段为云南的干季,降水占全年的15%左右,滇重楼植株容易受到干旱胁迫[23]。不同地区降水量的差异,导致干旱胁迫的程度不同,可能对滇重楼出苗及后期生长有不同影响。另外,研究发现土壤中氮、磷、钾、钙等常量元素和硼、钼、镧对重楼属植物生长发育和根茎生物量积累有一定程度的影响[24-25]。
本研究发现,滇重楼植株的根茎生物量均随生长年限增加而增加;相同生长年限植株中,元阳县样品根茎生物量变化范围大,且数值显著高于姚安县样品(<0.05)。该结果可能与滇重楼遗传多样性水平较高、居群间及居群内都具有一定的遗传分化等原因有关[26],是环境因子和遗传因素综合作用的结果[27]。陈铁柱等[28]研究发现华重楼根茎生物量随生长年限的增长,种植5年后是植株根茎生物量快速积累时期,从6年以后植株根茎生物量积累速度变慢。本研究与之不同,2个产地滇重楼种植5年后根茎生物量一直呈现快速积累趋势,与陈翠等[29-30]研究结果相似。
相同居群滇重楼植株的总生物量、株高、叶生物量、叶长、叶宽、叶面积随生长年限增加而增加,比叶面积则随生长年限增加而减小。不同居群滇重楼存在较大遗传差异性,叶片长度、叶片宽度、叶面积等叶片性状差异较大[31-32]。通过对2个产地同一年限的滇重楼植株比较可以发现,元阳县样品总生物量、叶生物量、叶长、叶宽、叶面积等指标显著高于姚安县样品(<0.05);元阳县样品株高平均值高于姚安县样品,但未达到显著差异;2个产地3~6年生样品根茎生物量占总生物量比例一致,但从第7年开始,元阳县样品根茎生物量/总生物量显著高于姚安县样品。本研究中,2个产地样品不同年限植株叶片数与比叶面积基本相同与李戈等[33]研究结果相似。
通过异速生长分析发现,与姚安县滇重楼样品相比,元阳县滇重楼样品根茎生物量积累速率高、株高较矮、叶生物量积累速率较低、比叶面积较大。提示为培育高产滇重楼,应考虑选择株高较矮、叶片较大而薄的地理种源。另外,环境资源的差异可以导致重楼属植物光合生理及形态特征发生改变,从而根茎生物量的积累[14, 34]。因此人工栽培滇重楼时需要注意管理,合理控制光照等因素,从而提高滇重楼药用部位产量。
利益冲突 所有作者均声明不存在利益冲突
[1] Whitman T, Aarssen L W. The leaf size/number trade-off in herbaceous angiosperms [J]., 2009, 3(1): 49-58.
[2] White J W, Montes-R C. Variation in parameters related to leaf thickness in common bean (L.) [J]., 2005, 91(1): 7-21.
[3] Yu K, Wang Y, Wei J R,. Improving rhizome yield and quality of Paristhrough gibberellic acid-induced retardation of senescence of aerial parts [J]., 2009, 4(5): 413-415.
[4] Shingleton A. Allometry: the study of biological scaling [J]., 2010, 3(10): 2.
[5] Weiner J. Allocation, plasticity and allometry in plants [J]., 2004, 6(4): 207-215.
[6] 张霁, 郭兰萍, 黄璐琦, 等. 异速生长及其在道地药材研究中的应用展望 [J]. 中国科学: 生命科学, 2013, 43(6): 457-463.
[7] Zhang J, Shen T, Zhao Y L,. The impact of human activity on the biomass allocation of a medicinal herbaceous species in an agroforestry system of Southwest China [J]., 2015, 89(3): 469-476.
[8] Zhang J, Zhao Y L, Jin H,. Morphological variability and allometric relationships of the herbin Yunnan, China [J]., 2017, 37(2): 65-69.
[9] 张霁, 赵艳丽, 王元忠. 野生和栽培滇龙胆草生物量分配及异速生长 [J]. 生态学杂志, 2018, 37(12): 3584-3589.
[10] 李恒. 重楼属植物 [M]. 北京: 科学出版社, 1998: 56.
[11] 中国药典 [S]. 一部. 2015: 260-260.
[12] Zhang J Y, Zhang J, Yang W Z,. Morphological diversity of wild medicinalL. from China and Vietnam [J]., 2011, 10(73): 16421-16428.
[13] 梁社往, 张世华, 何忠俊, 等. 不同地理种源滇重楼叶片碳氮代谢关键生理生化指标研究 [J]. 云南农业大学学报: 自然科学版, 2015(5): 771-776.
[14] 高成杰. 滇重楼生物量分配与环境调控机制研究 [D]. 北京: 中国林业科学研究院, 2015.
[15] 童凯, 孙旭, 姜美杰, 等. 华重楼的形态多样性及其与单株产量和质量的关系 [J]. 中国中药杂志, 2017, 42(7): 1300-1303.
[16] 李林玉, 李绍平, 杨斌, 等. 滇重楼不同居群形态变异研究 [J]. 中国中药杂志, 2010, 35(22): 2959-2962.
[17] 李绍平, 杨丽英, 杨斌, 等. 滇重楼高效繁育和高产栽培研究 [J]. 西南农业学报, 2008, 21(4): 956-959.
[18] Warton D I, Duursma R A, Falster D S,. Smatr 3- an R package for estimation and inference about allometric lines [J]., 2012, 3(2): 257-259.
[19] 侯秀丽, 赵峥, 王斌, 等. 滇重楼光合速率日变化及其对生态因子的响应 [J]. 江苏农业科学, 2015, 43(7): 265-267, 356.
[20] 苏文华, 张光飞. 滇重楼光合作用与环境因子的关系 [J]. 云南大学学报: 自然科学版, 2003, 25(6): 545-548.
[21] 刘涛, 王玲, 李玛, 等. 不同温度对滇重楼光合作用及有效成分含量的影响 [J]. 中国现代中药, 2015, 17(10): 1041-1043.
[22] 张家玲, 马英姿, 胡文俐, 等. 高温胁迫对滇重楼生理指标的影响 [J]. 中草药, 2018, 49(17): 4131-4137.
[23] 刘倩, 左应梅, 杨维泽, 等. 水分胁迫后复水对滇重楼生长以及皂苷积累的影响 [J]. 中药材, 2018, 41(10): 2277-2281.
[24] 何忠俊, 曾波, 韦建荣, 等. 钙对滇重楼生长和总皂甙含量的影响 [J]. 云南农业大学学报: 自然科学版, 2010, 25(5): 664-669.
[25] 牟杨. 几种常量元素(N、P、K)和微量元素(B、Mo、La)对华重楼生长及皂苷含量的影响 [D]. 雅安: 四川农业大学, 2016.
[26] 陈中苏直, 田波, 蔡传涛. 基于SSR分子标记的滇重楼遗传多样性研究 [J]. 中草药, 2017, 48(9): 1834-1838.
[27] 翁周, 王丽, 唐锐, 等. 多叶重楼的形态变异研究 [J]. 四川大学学报: 自然科学版, 2008, 45(5): 1228-1234.
[28] 陈铁柱, 文飞燕, 夏燕莉, 等. 生长年限对七叶一枝花植株、根系、产量和皂苷含量的影响 [J]. 江苏农业科学, 2018, 46(10): 122-125.
[29] 陈翠, 谭敬菊, 杨丽云, 等. 云南重楼生物量积累与营养分配研究 [J]. 西南农业学报, 2016, 29(2): 402-407.
[30] 陈翠, 张浩, 谭敬菊, 等. 云南重楼生长发育特性及皂苷含量动态变化研究 [J]. 西南农业学报, 2017, 30(6): 1320-1324.
[31] 陈翠, 康平德, 杨丽云, 等. 云南重楼不同地理种源分析评价与研究利用 [J]. 西南农业学报, 2009, 22(6): 1722-1726.
[32] 唐玲, 王艳芳, 李荣英, 等. 滇重楼不同居群的表型多样性研究 [J]. 中国农学通报, 2013, 29(18): 89-95.
[33] 李戈, 王艳芳, 唐玲, 等. 滇重楼表型性状变异、主成分、相关及通径分析 [J]. 中药材, 2015, 38(7): 1339-1342.
[34] 李剑美, 漆丽萍, 尚宇南, 等. 影响滇重楼幼苗农艺性状因子的研究 [J]. 普洱学院学报, 2016, 32(6): 1-5.
Biomass allocation and allometry ofvar.with different ages
WANG Yuan-zhong1, ZHANG Ji1, SHEN Tao2, ZHANG Jin-yu1
1. Medicinal Plants Research Institute, Yunnan Academy of Agricultural Sciences, Kunming 650200, China 2. College of Chemistry, Biology and Environment, Yuxi Normal University, Yuxi 653100, China
The differences of biomass and traits of cultivatedvar.in different geographic origins and growth years were compared. And allometry analysis was carried out on the law of biomass accumulation, growth and development in plants.Using Peareson correlation analysis to study the relevance between the rhizome biomass ofvar.and latitude, longitude, climate factors of collection site. One-way analysis of variance (One-Way ANOVA) and independent sample-test were used to compare the significant differences in plant traits between samples.The allometry of the biomass accumulation, growth and development ofwas studied by standardized major axis regression.Our research showed that the mean value of the rhizome biomass of the 5-year-old plants was 1.19—19.73 g, and there were significant differences among 12 origins (< 0.05), which did not significantly correlate with the longitude, latitude, mean annual temperature, and mean annual precipitation of the origins (>0.05). The rhizome biomass ofvar.increased with the growing age, and which even increased more rapidly after growing for five years. Among the same growing age plants, the samples of rhizome biomass from Yuanyang was significant higher (and coefficient of variation of the rhizome biomass) than the ones from Yao’an in Yunan Province (< 0.05). The total biomass, plant height, leaf biomass, leaf length, leaf width, and leaf area increased with growing age, whereas the specific leaf area (SLA) decreased with the growing age. The rhizome biomass/total biomass ratio increased with growing age in the samples from Yuanyang, while it didn’t change with growing age in the samples from Yao’an. Allometry relationships existed in rhizome biomass vs. total biomass, plant height vs. total biomass, leaf biomass vs. total biomass, and SLA vs. total biomass, respectively. Compared the samples from Yao’an, the samples from Yuanyang had higher accumulation rate for rhizome biomass, shorter height, lower accumulation rate for leaf biomass, and higher SLA.The results indicated that the geographical provenances with shorter height, large and thin leaves could be selected for maximizing thevar.yield. Furthermore, factors such as light should be considered when cultivatingvar.in order to increase the yield of the medicinal part of it.
var.(Franch.) Hand. - Mazz; biomass; allometry; functional trait; yield; growth
R286.2
A
0253 - 2670(2022)11 - 3456 - 07
10.7501/j.issn.0253-2670.2022.11.023
2021-12-06
国家自然科学基金项目(81460584);云南省科技计划项目(2017RA001)
王元忠(1981—),男,博士,副研究员,主要从事药用植物和真菌资源评价与利用。E-mail: boletus@126.com
张金渝,博士,研究员,主要从事植物资源评价与利用研究。E-mail: jyzhang2008@126.com
[责任编辑 时圣明]
猜你喜欢
农学学报(2022年8期)2022-11-16
飞天(2022年5期)2022-05-18
学与玩(2020年11期)2020-12-23
红楼梦学刊(2020年2期)2020-02-06
诗潮(2019年10期)2019-11-19
当代陕西(2019年6期)2019-04-17
当代县域经济(2017年3期)2017-03-06
农业与技术(2016年21期)2017-03-06
中国民族民间医药·上半月(2016年12期)2017-01-11
江苏农业科学(2016年9期)2016-11-28
